MITOSIS, MEIOSIS, CHROMOSOMES AND KARYOTYPES
OVERVIEW:
This lab has four sections. First, you will prepare and observe cell squashes of onion root tips
along with observing commercially prepared slides of fly salivary glands. You will see actual
chromosomes arrested in the stages of mitosis in the onion root, and you will see heterochromatic
and euchromatic bands on the polytene chromosomes found in fly salivary glands. This portion of
the lab should reinforce your understanding of what chromosomes are and how they move during
mitosis.
Second, you will conduct a quick mathematical exercise. You will use your understanding of the
meiotic process to prove your genetic individuality. Bring your calculator!
Third, we will begin to look at some Mendelian genetics. Using corn as an example, we will
observe a simple monohybrid cross.
Last, you will do a cut-and-paste exercise to familiarize yourself with karyotyping; a tool used by
clinical geneticists to screen for genetic abnormalities. Based on arm ratios and banding patterns,
you will sort chromosomes into homologous pairs, determine if there are any inconsistencies, and
provide a genetic diagnosis of the human subject.
Section I. Chromosomes and Mitosis
A. Polytene Chromosomes of the Drosophila melanogaster.
1. Introduction
The larval stage of flies is primarily concerned with food-gathering, and large salivary glands aid
in efficient digestion. These glands attain their large size by an increase in cell size, not an increase
in cell number (as in most other organs). Even after mitotic divisions have stopped, the
chromosomes continue to replicate as the cells enlarge. This produces thick chromosomes
consisting of thousands of sister chromatids in a parallel arrangement. (this extra DNA may aid the
oversized cell in satisfying its relatively large metabolic demand for enzymes and other proteins).
This thickened mass is much easier to see than a single chromosome. And, as all the strands in
these large polytene chromosomes are identical, they have the same areas of activity and banding.
These cells are locked in interphase permanently, and as a result, the chromosomes are
elongated and active. The multiplicative effect of replication allows us to see areas of activity that
would be impossible to distinguish in a single strand. These areas are bands. Areas that stain
darkly are areas of high DNA concentration; the DNA is wrapped up so tightly it is largely
nonfunctional. Light regions are areas where the DNA is diffuse; it is unraveled and active. When
these areas are transcribed, the DNA uncoils, 'puffing out'. This creates a chromosomal puff which
indicates active transcription. These are areas of highly concentrated RNA (as you might suspect!).
Additional anomalies also occur. First, the attractive forces of these strands is apparently very
great; so great that homologous chromosomes often fuse in parallel (like synapsis in Prophase I of
meiosis), thickening the strands even more. This effectively reduces the number of chromosomal
bodies by half, giving these cells a haploid chromosome number. In addition, the ends of the
chromosomes fuse at chromocenters. At this point, there is only one contorted chromosome; the
infamous GIANT CHROMOSOME! The study of giant chromosomes has been of tremendous
importance in genetics. Patterns of heredity and gene activity have been associated with specific
regions and bands along the chromosome. So, giant chromosomes have helped geneticists
understand how chromosome structure relates to function and gene expression.
2. Procedure
Observe the slides on demonstration on the front table. Answer the questions pertaining to the
polytene chromosomes at the end of this laboratory section.
B. Onion Root Tip Squash - Mitosis
1. Introduction
Root tips are meristematic regions; areas of rapid cell division. They are the perfect place to look
for cells proceeding through mitosis. During root development the meristematic region is located just
behind the root cap, a protective covering on the root. This can be seen as the bright white tip in the
preserved onion root tips we will use today in lab.
2. Procedures
a. Obtain two small cups, label one HCl, and place enough HCl in to cover the bottom.
b. Label the other cup 'Carnoy', and cover the bottom with Carnoy solution.
c. Transfer an onion root tip from the specimen bottle to your HCl cup. Leave for four minutes.
d. After four minutes, transfer the root tip to the Carnoy solution, and leave it for four minutes.
e. After four minutes, transfer the root tip to a slide. Cut off the distal 1-2 mm, and discard
all but this tip.
f. Cover the root tip in toluidine blue stain for two minutes.
g. After two minutes, blot away the stain; do not touch the root tip!
h. After blotting away the stain, cover the tip in water, and place a coverslip on the slide.
i. Press on the coverslip with an eraser; try not to twist the coverslip.
j. Find your cells under low power, then use greater magnification to search the tissue for
cells in the various stages of mitosis. Recognize that the process is continuous; the
discrete stages we have discussed are connected by intermediate steps. Draw
pictures of the stages you see, labeling them as appropriate. During telophase,
notice cell wall deposition.
Section II. Your Genetic Individuality
Consider this: a diploid organism with 4 chromosomes (2n=4) can produce four different gametes
(22), since chromosome 'A' of the first homologous pair is not identical to 'a' and 'B' is not identical to 'b'.
1st Homologous Pair = A a
2nd Homologous Pair = B b
Gametes: AB, Ab, aB, ab
An organism with 2n=6 can produce 8 gametes (23).
Extrapolate to an organism with 2n=46, such as Homo sapiens.
How many different kinds of gametes can one person produce?
Now, how many different zygotes can two humans produce? This is actually a gross underestimation
of your uniqueness, since we haven't even considered recombination (crossing over)! Even so, this
number is far in excess of the number of people who have ever lived, and so it is quite likely that you
are 'one-of-a-kind' (but you knew that already, didn't you).
Section III: The Monohybrid Cross
1. Introduction:
Gregor Mendel, a nineteenth-century Austrian monk, devised a model that predicted patterns of
inheritance. a report of his meticulous records, first printed in 1866, was discovered in 1900. The
elements of heredity that Mendel called "factors" are now known to be genes. Even though Mendel
did not know of chromosomes or genes, his work opened the door to an understanding of heredity.
Mendel conducted his experiments with garden peas in three phases. First he established a true-
breeding population. In this population of plants the offspring (F1, the First Filial Generation) carried
the same characteristics as the parents.(P1). Mendel then crossed varieties of peas expressing
alternative traits. For example, he crossed peas that were tall with peas that were dwarf. The
offspring (F1) of these crosses would only express one of the two alternative traits. That trait which
was expressed in the F1 generation was termed "dominant" while the alternative trait was termed
"recessive". Finally, Mendel allowed the F1 offspring to self-pollinate (cross among themselves), and
he counted and carefully recorded the number of each characteristic in the progeny. For the first
time, numerical data pertaining to inheritance could be analyzed. Mendel discovered that the F2
generation (second filial generation) expressed alternative traits in a ratio of three of one trait to one
of the alternate trait (3:1). This 3:1 ratio is a phenotype ratio. The phenotype of an organism is the
observable features of an individual organism that result from an interaction between the genotype
and the environment in which development occurs. The genotype is the genetic constitution of an
individual, usually referring to specific characters under consideration. Thus, the two alternative
alleles for the tall and dwarf genes of the pea plants could be written D and d. A capital letter is used
to designate dominance while a lower case letter is used to designate the recessive trait. Mendel
noted that traits always exist in pairs, thus for a plant showing the tall phenotype there are two
possible genotypes DD and Dd. For a plant showing the dwarf phenotype there is only one
genotype dd.
2. Procedure:
Observe the ears of corn. Note that each kernel of corn is a seed that can establish an
individual plant. Thus, by counting the individual corn kernels, you are really counting individual
plants. Note the different characteristics of the kernels. These are alternative traits, purple and
yellow kernels, follow Mendel�s observations. These ears of corn where obtained by first crossing P1
that were all purple kernel (YY) plants with all yellow kernel (yy) plants. The resulting F1 generation
was then used to produce the corn you are observing. Use these ears of corn to answer the
questions at the end of this laboratory exercise.
Section IV. Human Karyotype Analysis
Cytogenetics, the study of chromosomes, has its origin more than a century ago. However, not until
the last 3O years or so have the human chromosome studies become a major field in the biomedical
sciences. Chromosome banding methods, for example, are today's vital tools in clinical genetics and
evolutionary studies. Furthermore, human cytogenetics coupled with molecular techniques has
revolutionized the field of molecular genetics, including gene mapping and recombinant DNA
technology.
Prior to 1956, the number of human chromosomes was estimated to be between 37 and 48. The
difficulty of determining the exact chromosome number was due to the lack of proper techniques for
cell culture and for spreading the chromosomes for microscopic observations. In the 1950's two
major breakthroughs led to the rapid advancement in human cytogenetics: (1) the use of
phytohemaglutinin (PHA), a substance from plants, to stimulate cell division of lymphocytes (white
blood cells), so that a large amount of cells in metaphase could be obtained, and (2) the use of
hypotonic (low salt) solution to cause cells to absorb water and swell, resulting in spreading and
separating the individual chromosomes, so that they were readily distinguishable. In 1956,
researchers in Sweden systematically counted 46 chromosomes in human cells. Three years later,
the discovery of an extra chromosome 21 in patients with Down syndrome marked the beginning of
clinical application of chromosome studies, and soon the identification of other chromosome
disorders followed.
Conventional dyes such as aceto-orcein used for direct chromosome staining could only distinguish
chromosomes according to their sizes and centromere positions. On the basis of these two criteria,
human chromosomes were classified into seven groups, A to G. Accurate chromosome identification
started in the late 1960's when quinocrine mustard (QM) was used to stain human chromosomes.
With this technique, chromosomes were differentiated into bright and dark regions, called Q-bands.
Since then, numerous banding techniques have been developed. Of these, G-banding is the most
widely used technique for chromosome analysis. The banding techniques are extremely useful for
the detection of structural changes. Clinical applications include the association of chromosome
disorders with numerous diseases, including cancers.
Today chromosome analysis of cultured amniotic fluid cells is routinely performed in medical
genetics laboratories for prenatal diagnosis. For structural changes which are too small to be
detected by direct microscope examinations, other approaches, such as the use of known DNA
sequence (DNA probe) to hybridize the homologous DNA of the chromosome (in situ hybridization),
are available. The integration of cytogenetics with molecular technology has thus opened up an
exciting field in modem biology.
Chromosome Morphology
1. The Cell Cycle
Chromosomes are not visible under the light microscope in non-dividing (interphase) cells. As the
cell begins to divide, the threads of chromatin (DNA-protein complex) in the nucleus start to
condense into multiple levels of coiled structure recognizable as chromosomes. There are two types
of cell division: mitosis and meiosis. Mitosis is responsible for the proliferation of body (somatic)
cells, whereas meiosis is responsible for the production of gametes. Because mitotic cells are easy
to obtain, morphological studies are generally based on mitotic metaphase chromosomes.
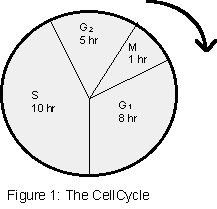
The transition of interphase to cell division (mitosis) and back to interphase is called the cell
cycle. For a dividing cell the interphase can be divided into three stages: G1, S, and G2. (G means
gap.) During the G1 stage, cytoplasmic components such as membranes, organelles, and ribosomes
begin to proliferate. G1 is followed by S stage during which DNA synthesis occurs so that the
amount of DNA per chromosome is doubled, resulting in two sister-chromatids visible during
prophase and metaphase. S is followed by a second period of growth, the G2 stage. At the end of
the G2 stage, the cell starts to divide, thus entering the M (mitosis) stage. When the cell is not
cycling, the interphase is said to be in Go. The cell cycle for human cells averages about 24 hours (8
h for G1, 10 h for S, 5 h for G2, and 1 h for M).
Mitosis can be divided into four phases: prophase, metaphase, anaphase, and telophase.
During prophase, chromosomes, consisting of two chromatids each, condense and undergo
progressive coiling. In the early prophase they appear as thin threads. At the middle of late
prophase the chromosomes can be seen as discrete structures, each with two chromatids and a
constricted area called the centromere where the two chromatids are joined. The transitional period
from late prophase to metaphase is called prometaphase, during which the nuclear envelope breaks
down and the chromosomes move toward the equatorial plane (metaphase plate) of the cell.
2. Chromosome Structure
At metaphase, the chromosomes are at the most condensed state with spindle fibers attaching to
the area of the centromere called kinetochore, forming pole-chromosome fibers. The fibers which
bypass the centromeres are called pole-pole fibers. Anaphase begins with the division of the
centromere and the separation of chromatids. Once separate, each chromatid is now known as a
chromosome. Telophase marks the final stage of mitosis, during, which the cell undergoes a reverse
process of prophase, including chromosome uncoiling, spindle fiber disappearing, and nuclear
envelope and nucleolus reappearing. At the same time, the cytoplasm is divided (cytokinesis) into
two parts, forming two daughter cells.
The best mitotic stage for chromosome analysis is prometaphase or metaphase. A typical
metaphase chromosome consists of two arms separated by a primary constriction or centromere.
Each of the two sister-chromatids contains a highly coiled double helix of DNA. A chromosome may
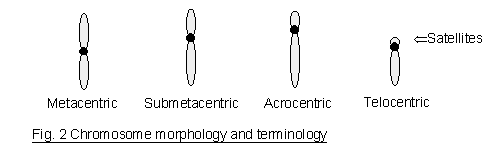
be characterized by its total length and the position of its centromere (Fig. 2). A chromosome with
the centromere at or near the middle is called metacentric. A submetacentric chromosome is
referred to as the one with a centromere somewhat off the middle point. If the centromere is
obviously off center (e.g., halfway between the middle and the tip of the chromosome), the term
acrocentric may be used. Telocentric chromosomes, which are absent in human cells, have the
centromere at the very tip of one end. The short chromosome arm is designated p (petite) and the
long arm q (one letter after p). Certain human chromosomes may also contain a secondary
constriction, which appears as an unstained gap (also called a satellite stalk) near the tip. The
chromosomal segment distal to this gap appears as a satellite. Chromosome numbers 13-15 (D
group) and 21-22 (G group) have satellites.
 3. Chromosome Number
The diploid chromosome number is the number of chromosomes in the somatic cell and is
designated by the symbol 2N. The gametes, which have one-half the diploid number, have haploid
number N. In humans, the diploid number is 46, with 23 from each parent through the sperm or the
egg. Same (homologous) chromosomes form a pair with one member from each parent. Thus, there
are 23 pairs of chromosomes in human cells. Of these, 22 pairs are not directly involved in sex
determination, known as autosomes. The remaining chromosome pair which is directly involved in
sex determination is called the sex chromosome. In females the two sex chromosomes are identical
(XX); whereas in males the two sex chromosomes are not identical (XY). The Y chromosome is
smaller than the X chromosome. A gene which is responsible for switching the embryo to male
characteristics was recently discovered in the Y chromosome.
4. G-banding
The purpose of banding techniques is to uniquely identify chromosomes or portions of
chromosomes, both normal and abnormal. Most laboratories in this country use a form of Giemsa
banding (G-banding) as a general purpose banding method. Giemsa reagent is a DNA stain which
consists of a mixture of dyes including the basic aminophenothiazine dyes (azure A, azure B, azure
C, thionin, and methylene blue) and the acidic dye (eosin). Prior to staining, some form of protease
(an enzyme which digests proteins) treatment is needed. This technique produces patterns of light-
staining (G-light) regions and dark-staining (G-dark) regions. The pattern is consistent to each
chromosome, and, therefore, it serves as a landmark for chromosome identification.
The G-light bands are thought to be relatively GC-rich, and the G-dark bands relatively AT-rich.
Furthermore, the light bands represent the regions which are relatively open and which contain most
of the genes including housekeeping genes (genes active in every cell type). On the other hand, the
G-dark bands represent regions which are relatively compact and contain few genes. The genes in
the dark regions are mainly tissue specific.
It should be noted that the pattern of light and dark bands changes with the stage in mitosis,
ranging from 300 or less in late metaphase to more than 1000 bands in early prophase. As mitosis
progresses, bands coalesce, accounting for the reduction of bands. Most routine lab work is done at
350-400 band level. For critical analysis, special techniques must be employed to produce 550 or
more bands, known as high resolution analysis.
5. Karyotype
The term karyotype refers to a display of the chromosomes of a cell by lining them up,
beginning with the largest and with the short arm oriented toward the top of the karyotyping
sheet. A diagram of karyotype based on chromosome measurements in many cells is called an
ideogram.
6. Nomenclature
Human chromosome nomenclature systems were developed in six international conferences on
chromosome nomenclature. Two systems, one short and the other detailed, are currently used by
cytogenetists. For writing a karyotype description, work from left to right and leave no spaces.
Separate each item with a comma unless otherwise specified. The karyotype formula begins with the
total number of chromosomes in the cell which is followed by the sex chromosomes (first the X's and
then the 'Ys). For example, the formula for a normal male is "46,XY" and for a normal female is
"46,XX". An extra or a missing chromosome is designated with a "+" or "-" sign, respectively, before
the number of chromosome. Thus, a male with trisomy (three chromosomes) for chromosome 18 is
47, XY, + 18, and a female with a monosomy (one chromosome) for 22 is 45, XX -22. Addition or
deletion of a chromosome segment is denoted with a plus "+" or "-" sign after the symbol of the
chromosome arm, respectively. For example, the formula for a female with the cri du chat syndrome
(resemblance of the cry of an affected newborn to cat mewing) is 46, XX, 5p-(missing a piece of
short arm of chromosome 5). The formula for a male with translocation (exchanging chromosome
segments) between chromosome 14 and 21 is 46, XY, t(14-21).
The following is a partial list of symbols for chromosome nomenclature.
A-G The chromosome groups
1-22 The autosome numbers
X,Y The sex chromosomes
del Deletion
der Derivative of chromosome
dup Duplication
ins Insertion
inv Inversion
p Short arm of chromosome
q Long arm of chromosome
s Satellite
t Translocation
+ or - Placed before the chromosome number, they indicate addition (+) or loss (-) of the
whole chromosome; e.g. +21 denotes an extra chromosome 21, as in Down
syndrome. Placed after the chromosome number, these symbols indicate
increase or decrease in the length of the chromosome part; e.g., 5p- indicates
loss of part of the short arm of chromosome 5, as in cri du chat syndrome.
Objectives:
1. To learn the human chromosome classification system
2. To use G-banding landmarks to identify individual chromosomes
3. To learn the karyotyping procedure
Procedure:
1. First, familiarize yourself with such terms as metacentric, submetacentric, very submetacentric,
acrocentric; p arm, q arm; proximal and distal regions of a chromosome arm; and satellite.
Then study the banding pattern of each chromosome as well as the grouping of chromosomes
from A to G (based on size, X chromosomes belong to C group and Y chromosomes belong to
G group). Note that groups D and G are the only two groups having acrocentric
chromosomes with satellites.
2. Count the total number of chromosomes on your work sheet (Metaphase Spread).
3. Use scissors to cut out each chromosome from the Metaphase Spread.
4. Arrange all individual chromosomes according to size, with the longest one first, on a smooth
surface such as the clean lab bench.
5. Group similar chromosomes as a pair if possible.
6. Read the description of the Human Chromosomes (below) and arrange each pair of
chromosomes.
7. When some of the chromosomes are identified, place them on the Karyotype Form. Critically
identify the location of the centromers for each chromosome and be sure that the p arm is up
and the q arm is down; upside down chromosomes are frequent sources of misidentification.
8. Check with your instructor for accuracy. When all the chromosomes are correctly identified,
arrange on the karyotype form. Use a glue stick to glue each chromosome picture firmly on
the Karyotype Form.
9. Write on the top of the Worksheet the karyotype formula of your sample.
Human Chromosome Descriptions
Chromosome 1 (Group A)
The longest chromosome, metacentric; p arm fading at distal end, with least 2 dark bands above
centromere; q arm with a dark area just below centromere, a light band below, followed by 4 dark
bands of which the middle 2 are often more intense.
Chromosome 2 (Group A)
Almost as long as chromosome 1, submetacentric; p arm without light distal end, with 4 dark bands;
q arm with more than 4 dark bands.
Chromosome 3 (Group A)
Metacentric; both p and q arms with a large light band in the middle; dark area (consisting of 2 or 3
close dark bands) distal to the light band of q arm thicker than its counterpart in p arm.
Chromosome 4 (Group B)
Very submetacentric, p arm with a thick dark band which may sometimes be resolved as double; q
arm with a distinct, prominent dark band below centromere.
Chromosome 5 (Group B)
Very submetacentric; p arm with a single dark band; q arm without a prominent dark band below
centromere, with 3 close dark bands in the middle.
Chromosome 6 (Group C)
Submetacentric; p arm with a distinct, large light band in the middle; q arm with several dark bands.
Chromosome 7 (Group C)
Submetacentric; p arm with a prominent flat dark band at the top; q arm with 2 distinct dark bands
followed by a much less intense dark band.
Chromosome 8 (Group C)
Very submetacentric; p arm with 2 dark bands (sometimes unclear); q arm with 2 (sometimes 3)
dark bands, the distal dark band characteristically more intense.
Chromosome 9 (Group C)
Varying from slight submetacentric to very submetacentric; p arm with a broad (often square in shape
) dark band (which may be resolved as double); q arm with a variable, proximal light area (from as
long as the p arm to almost non-existent) followed by two distinct, equally intense broad dark bands
with a light band between them, these.' ) bands (2 dark bands and one light band) equal in thickness.
Chromosome 10 (Group C)
Submetacentric; p arm with a dark band; q arm with 3 distinct dark bands, the proximal dark band
most intense.
Chromosome 11 (Group C)
Submetacentric; p arm with I or occasionally 2 dark bands; q arm with a very large light area followed
by a dark band which may be resolved into 2 dark bands.
Chromosome 12 (Group C)
Very submetacentric; p arm with a dark band; q arm with a proximal light band (which is relatively
thinner than the counterpart of chromosome I 1) followed by a dark area in the middle which may be
resolved into 2, or more often 3, bands.
Chromosome 13 (Group D)
Acrocentric- p arm with satellites (visible or invisible); q arm with several dark bands.
Chromosome 14 (Group D)
Acrocentric; p arm with satellites (visible or invisible); q arm with a distal dark band which is more
intense than other dark bands.
Chromosome 15 (Group D)
Acrocentric; p arm with satellites (visible or invisible); q arm with dark proximal half and light distal
half
Chromosome 16 (Group E)
Metacentric or occasionally submetacentric; p arm light or slightly dark; q arm with a prominent dark
band just below centromere, followed by I or 2 lesser dark bands.
Chromosome 17 (Group E)
Submetacentric or very submetacentric; p arm with a thin dark band (may be invisible); centromere
dark followed by a distinct, large light band occupying most part of q arm, q arm with I or 2 distinct
dark bands near the terminal end.
Chromosome 18 (Group E)
Submetacentric or very submetacentric; p arm often uniformly dark; q arm with 2 distinct dark bands,
one proximal and one distal; the light band between these 2 dark bands slightly smaller than the
counterpart of chromosome 17.
Chromosome 19 (Group F)
Metacentric; p and q arms both light; centromere dark.
Chromosome 20 (Group F)
Metacentric; p arm dark; q arm light, with 2 narrow, less intense dark bands.
Chromosome 21 (Group G)
Acrocentric; p arm with satellites (visible or invisible); q arm with intense dark area proximally, fading
out toward distal end.
Chromosome 22 (Group G)
Acrocentric; p arm with satellites (visible or invisible); centromere dark; q arm with a narrow, less
intense dark band.
X Chromosome (sex chromosome)
Submetacentric; p arm with distinct, strong dark at the middle; q arm with a proximal dark band about
the same distance from the centromere as the p arm dark band, with two less intense dark bands
near distal end.
Y Chromosome (sex chromosome)
Very submetacentric; p arm very short or invisible; q arm dark throughout.
Section IV. Wrap-Up
Make sure your station is clear and equipment is away. Hand in your exercise sheet. I'll return
your drawings for study purposes (you just never know when this stuff might pop up; its fair game
come exam time!)
Click here to dispay the Student Worksheet needed during the lab period
3. Chromosome Number
The diploid chromosome number is the number of chromosomes in the somatic cell and is
designated by the symbol 2N. The gametes, which have one-half the diploid number, have haploid
number N. In humans, the diploid number is 46, with 23 from each parent through the sperm or the
egg. Same (homologous) chromosomes form a pair with one member from each parent. Thus, there
are 23 pairs of chromosomes in human cells. Of these, 22 pairs are not directly involved in sex
determination, known as autosomes. The remaining chromosome pair which is directly involved in
sex determination is called the sex chromosome. In females the two sex chromosomes are identical
(XX); whereas in males the two sex chromosomes are not identical (XY). The Y chromosome is
smaller than the X chromosome. A gene which is responsible for switching the embryo to male
characteristics was recently discovered in the Y chromosome.
4. G-banding
The purpose of banding techniques is to uniquely identify chromosomes or portions of
chromosomes, both normal and abnormal. Most laboratories in this country use a form of Giemsa
banding (G-banding) as a general purpose banding method. Giemsa reagent is a DNA stain which
consists of a mixture of dyes including the basic aminophenothiazine dyes (azure A, azure B, azure
C, thionin, and methylene blue) and the acidic dye (eosin). Prior to staining, some form of protease
(an enzyme which digests proteins) treatment is needed. This technique produces patterns of light-
staining (G-light) regions and dark-staining (G-dark) regions. The pattern is consistent to each
chromosome, and, therefore, it serves as a landmark for chromosome identification.
The G-light bands are thought to be relatively GC-rich, and the G-dark bands relatively AT-rich.
Furthermore, the light bands represent the regions which are relatively open and which contain most
of the genes including housekeeping genes (genes active in every cell type). On the other hand, the
G-dark bands represent regions which are relatively compact and contain few genes. The genes in
the dark regions are mainly tissue specific.
It should be noted that the pattern of light and dark bands changes with the stage in mitosis,
ranging from 300 or less in late metaphase to more than 1000 bands in early prophase. As mitosis
progresses, bands coalesce, accounting for the reduction of bands. Most routine lab work is done at
350-400 band level. For critical analysis, special techniques must be employed to produce 550 or
more bands, known as high resolution analysis.
5. Karyotype
The term karyotype refers to a display of the chromosomes of a cell by lining them up,
beginning with the largest and with the short arm oriented toward the top of the karyotyping
sheet. A diagram of karyotype based on chromosome measurements in many cells is called an
ideogram.
6. Nomenclature
Human chromosome nomenclature systems were developed in six international conferences on
chromosome nomenclature. Two systems, one short and the other detailed, are currently used by
cytogenetists. For writing a karyotype description, work from left to right and leave no spaces.
Separate each item with a comma unless otherwise specified. The karyotype formula begins with the
total number of chromosomes in the cell which is followed by the sex chromosomes (first the X's and
then the 'Ys). For example, the formula for a normal male is "46,XY" and for a normal female is
"46,XX". An extra or a missing chromosome is designated with a "+" or "-" sign, respectively, before
the number of chromosome. Thus, a male with trisomy (three chromosomes) for chromosome 18 is
47, XY, + 18, and a female with a monosomy (one chromosome) for 22 is 45, XX -22. Addition or
deletion of a chromosome segment is denoted with a plus "+" or "-" sign after the symbol of the
chromosome arm, respectively. For example, the formula for a female with the cri du chat syndrome
(resemblance of the cry of an affected newborn to cat mewing) is 46, XX, 5p-(missing a piece of
short arm of chromosome 5). The formula for a male with translocation (exchanging chromosome
segments) between chromosome 14 and 21 is 46, XY, t(14-21).
The following is a partial list of symbols for chromosome nomenclature.
A-G The chromosome groups
1-22 The autosome numbers
X,Y The sex chromosomes
del Deletion
der Derivative of chromosome
dup Duplication
ins Insertion
inv Inversion
p Short arm of chromosome
q Long arm of chromosome
s Satellite
t Translocation
+ or - Placed before the chromosome number, they indicate addition (+) or loss (-) of the
whole chromosome; e.g. +21 denotes an extra chromosome 21, as in Down
syndrome. Placed after the chromosome number, these symbols indicate
increase or decrease in the length of the chromosome part; e.g., 5p- indicates
loss of part of the short arm of chromosome 5, as in cri du chat syndrome.
Objectives:
1. To learn the human chromosome classification system
2. To use G-banding landmarks to identify individual chromosomes
3. To learn the karyotyping procedure
Procedure:
1. First, familiarize yourself with such terms as metacentric, submetacentric, very submetacentric,
acrocentric; p arm, q arm; proximal and distal regions of a chromosome arm; and satellite.
Then study the banding pattern of each chromosome as well as the grouping of chromosomes
from A to G (based on size, X chromosomes belong to C group and Y chromosomes belong to
G group). Note that groups D and G are the only two groups having acrocentric
chromosomes with satellites.
2. Count the total number of chromosomes on your work sheet (Metaphase Spread).
3. Use scissors to cut out each chromosome from the Metaphase Spread.
4. Arrange all individual chromosomes according to size, with the longest one first, on a smooth
surface such as the clean lab bench.
5. Group similar chromosomes as a pair if possible.
6. Read the description of the Human Chromosomes (below) and arrange each pair of
chromosomes.
7. When some of the chromosomes are identified, place them on the Karyotype Form. Critically
identify the location of the centromers for each chromosome and be sure that the p arm is up
and the q arm is down; upside down chromosomes are frequent sources of misidentification.
8. Check with your instructor for accuracy. When all the chromosomes are correctly identified,
arrange on the karyotype form. Use a glue stick to glue each chromosome picture firmly on
the Karyotype Form.
9. Write on the top of the Worksheet the karyotype formula of your sample.
Human Chromosome Descriptions
Chromosome 1 (Group A)
The longest chromosome, metacentric; p arm fading at distal end, with least 2 dark bands above
centromere; q arm with a dark area just below centromere, a light band below, followed by 4 dark
bands of which the middle 2 are often more intense.
Chromosome 2 (Group A)
Almost as long as chromosome 1, submetacentric; p arm without light distal end, with 4 dark bands;
q arm with more than 4 dark bands.
Chromosome 3 (Group A)
Metacentric; both p and q arms with a large light band in the middle; dark area (consisting of 2 or 3
close dark bands) distal to the light band of q arm thicker than its counterpart in p arm.
Chromosome 4 (Group B)
Very submetacentric, p arm with a thick dark band which may sometimes be resolved as double; q
arm with a distinct, prominent dark band below centromere.
Chromosome 5 (Group B)
Very submetacentric; p arm with a single dark band; q arm without a prominent dark band below
centromere, with 3 close dark bands in the middle.
Chromosome 6 (Group C)
Submetacentric; p arm with a distinct, large light band in the middle; q arm with several dark bands.
Chromosome 7 (Group C)
Submetacentric; p arm with a prominent flat dark band at the top; q arm with 2 distinct dark bands
followed by a much less intense dark band.
Chromosome 8 (Group C)
Very submetacentric; p arm with 2 dark bands (sometimes unclear); q arm with 2 (sometimes 3)
dark bands, the distal dark band characteristically more intense.
Chromosome 9 (Group C)
Varying from slight submetacentric to very submetacentric; p arm with a broad (often square in shape
) dark band (which may be resolved as double); q arm with a variable, proximal light area (from as
long as the p arm to almost non-existent) followed by two distinct, equally intense broad dark bands
with a light band between them, these.' ) bands (2 dark bands and one light band) equal in thickness.
Chromosome 10 (Group C)
Submetacentric; p arm with a dark band; q arm with 3 distinct dark bands, the proximal dark band
most intense.
Chromosome 11 (Group C)
Submetacentric; p arm with I or occasionally 2 dark bands; q arm with a very large light area followed
by a dark band which may be resolved into 2 dark bands.
Chromosome 12 (Group C)
Very submetacentric; p arm with a dark band; q arm with a proximal light band (which is relatively
thinner than the counterpart of chromosome I 1) followed by a dark area in the middle which may be
resolved into 2, or more often 3, bands.
Chromosome 13 (Group D)
Acrocentric- p arm with satellites (visible or invisible); q arm with several dark bands.
Chromosome 14 (Group D)
Acrocentric; p arm with satellites (visible or invisible); q arm with a distal dark band which is more
intense than other dark bands.
Chromosome 15 (Group D)
Acrocentric; p arm with satellites (visible or invisible); q arm with dark proximal half and light distal
half
Chromosome 16 (Group E)
Metacentric or occasionally submetacentric; p arm light or slightly dark; q arm with a prominent dark
band just below centromere, followed by I or 2 lesser dark bands.
Chromosome 17 (Group E)
Submetacentric or very submetacentric; p arm with a thin dark band (may be invisible); centromere
dark followed by a distinct, large light band occupying most part of q arm, q arm with I or 2 distinct
dark bands near the terminal end.
Chromosome 18 (Group E)
Submetacentric or very submetacentric; p arm often uniformly dark; q arm with 2 distinct dark bands,
one proximal and one distal; the light band between these 2 dark bands slightly smaller than the
counterpart of chromosome 17.
Chromosome 19 (Group F)
Metacentric; p and q arms both light; centromere dark.
Chromosome 20 (Group F)
Metacentric; p arm dark; q arm light, with 2 narrow, less intense dark bands.
Chromosome 21 (Group G)
Acrocentric; p arm with satellites (visible or invisible); q arm with intense dark area proximally, fading
out toward distal end.
Chromosome 22 (Group G)
Acrocentric; p arm with satellites (visible or invisible); centromere dark; q arm with a narrow, less
intense dark band.
X Chromosome (sex chromosome)
Submetacentric; p arm with distinct, strong dark at the middle; q arm with a proximal dark band about
the same distance from the centromere as the p arm dark band, with two less intense dark bands
near distal end.
Y Chromosome (sex chromosome)
Very submetacentric; p arm very short or invisible; q arm dark throughout.
Section IV. Wrap-Up
Make sure your station is clear and equipment is away. Hand in your exercise sheet. I'll return
your drawings for study purposes (you just never know when this stuff might pop up; its fair game
come exam time!)
Click here to dispay the Student Worksheet needed during the lab period
You will need the Student Worksheet and the Karyotype Worksheet (found as a link at the end of the
Student Worksheet) for the laboratory period.