Transport in the
Phloem
Transport of Organic Solutes
-
Girdling Experiments:
-
Transpiration stream occurs in the wood -
xylem
-
Nutrients essential to life move in the bark
- phloem
-
Assimilates: products of assimilation
& metabolism
-
Inorganic ions (phosphate, sulfate,
potassium) remain chemically unchanged during transport. Sometimes
phosphate & sulfate are incorporated into nucleotides & amino acids.
-
CO2 is first incorporated into
sucrose or some other sugar & is then eventually incorporated into
every organic compound in the plant
The Pressure-Flow Mechanism
-
Study how the laboratory system with the two
osmometers is set up and how it works. This system shows how bulk
flow occurs in the phloem of plants.
-
Bulk flow (mass flow).
-
Flow through the sieve tubes is passive, occurring
in response to the pressure gradient caused by osmotic diffusion of water
into sieve tubes at the source end of the system & out of the sieve
tubes at the sink end of the system.
-
Cytoplasmic streaming does not occur in mature
sieve elements & thus cannot play a role in phloem transport.
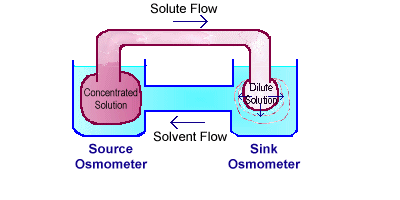
A lab model showing two
osmometers that illustrates the pressure-flow theory of solute translocation
in phloem
Testing the Hypothesis
-
Phloem anatomy
-
Phloem Tissue
-
Sieve elements or sieve tube members (no nuclei)
-
Angiosperms: sieve plates between cells.
Forms sieve tubes.
-
Gymnosperms: sieve plates are not as
clearly displayed., sieve cells
-
Companion cells (Angiosperms) -- Albuminous
cells (Gymnosperms) (nuclei, with copious plasmodesmata, exact function
not known)
-
Phloem parenchyma: storage & lateral
transport
-
Phloem fibers: support
-
Minor & Major leaf veins. No mesophyll
cell in leaf is separated from a minor vein by more than 2 or 3 other mesophyll
cells.
-
Transfer Cells:
-
Phloem Development
-
Slime Bodies: phloem
protein (P-protein)
-
Phloem Ultrastructure:
-
Rates of Phloem Transport
-
Mass transfer rate: quantity of material
passing through a given cross section of sieve tubes per unit of time.
-
Velocity: linear distance traversed
by an assimilate molecule per unit of time.
-
Osmotic potentials of -2 to -3 MPa are common
in intact sieve tubes. Equivalent to 20 - 30 % sucrose solution.
-
Transported Solutes
-
Bleeding is often rapidly stopped as P-protein
& other particulate matter clog the sieve pores.
-
90 % or more of the material transloacted
in phloem consists of carbohydrates.
-
Inorganic nitrogen is transported in the xylem
as nitrate (NO3-), which is not present in phloem sap. Nitrogen can
be transported in xylem as ureides, amides, or other nitrogen-rich molecules.
-
Nutritional completeness of sieve-tube sap.
Many plant parts with no or minimal transpiration are completely dependent
on the phloem for organic & inorganic nutrients during part or all
of their growth.
-
Phloem Loading: Mesophyll cells have
osmotic potentials of -1.3 to -1.8 MPa. Sieve elements in leaves
have osmotic potentials of -2.0 to -30. MPa. Osmotic potential is
caused by presence of sugars. Sugar concentration is approximately
1.5 - 3 times higher in sieve elements than surrounding mesophyll cells.
-
Pathway of transport
-
Sugar is actively secreted from mesophyll
cells into apoplast of minor veins. Sugar then adsorbed actively
into large companion cells of minor veins. Then passes symplastically
into sieve elements.
-
Phloem loading can sometimes occur via symplast.
-
Some species may use element of both symplast
& apoplast paths
-
Active loading of sucrose into companion cells
could produce a strongly negative osmotic potential in those cells, leading
to an osmotic entrance of water, which would then pass in bulk flow across
the plasmodesmatal connections between the companion cells & sieve
element, carrying the sucrose along with it. High pressures &
mass flow.
-
Selective loading of sugars
-
Only sugars that are transported in phloem
are accumulated in minor veins. Selectivity of phloem loading is
based on mutual recognition by sugars & carriers in the plasmalemma.
-
Amino acids are preferentially loaded.
Minerals also show variable loading capacities
-
Many substances can enter phloem passively
by diffusing in along their own concentration gradients. (growth regulators)
-
Sucrose/proton cotransport mechanism:
transport of organic molecules such as sugars & amino acids is linked
with transport of hydrogen ions.
-
Role of metabolism in transport: maintenance
of phloem transport system for bulk flow of sap apparently requires only
a minimum of metabolic energy. Metabolic energy is required for phloem
loading.
-
Development of loading capacity: development
of phloem loading capacity in minor veins could account for switch from
import to export. Once sucrose is actively loaded into sieve elements,
water will enter by osmosis, & flow will begin out of the minor veins;
leaf becomes a source instead of a sink.
-
Phloem Unloading: maintains low
phloem turgor pressures at the sink. Solute unloaded at sink can
then be absorbed into developing fruit or other cells. Concentrations
can reach values higher than in sieve tubes at source.
-
Pressure in the Phloem: a pressure gradient
occurs in phloem sufficient to account for flow from source to sink.
Gradients in osmotic potential in sieve tubes from source to sink have
often been measured, with most negative values at source.
-
Two Problems with Pressure Flow: Substances
should move in the phloem not only in the same direction but at the same
velocity. Studies seem to contradict this showing that water moved
slower than solutes. What accounts for this problem? (see the
2 explanations)
-
Pressure Flow: Summary: Munch
osmometer experiment reviewed. Osmotic system (symplast) with surrounding
membranes exists in the plant, & pressures are observed in the transport
system. The medium with high water potential around the source phloem
tissues is the hydrated apoplast.
Partitioning & Control Mechanisms
-
Photosynthesis & Sink Demand
-
Photosynthesis of leaves is strongly
influenced by sink demands
-
When sink demand is low, sucrose piles up
on the leaves, causing a product inhibition of photosynthetic reactions.
-
Metabolically Driven Gradients
-
Sucrose gradient that drives phloem transport
is produced by metabolism of sucrose in sink tissues.
-
In storage tissues sucrose may be changed
to starch or some other less-soluble product that has a lower effect on
osmosis than sucrose.
-
Phloem Unloading
-
Removal of sucrose or other solutes from sieve
cells makes the osmotic potential in those sieve cells less negative, so
pressure transmitted from the source areas raises the water potential even
further, & water diffuses out into the apoplast.
-
Decreasing pressure in the sink end of the
sieve tubes increases the pressure gradient between source & sink &
leads to further flow toward the sink region.
-
Growth Regulator Directed Transport
-
Growth regulators help direct translocation.
-
New sinks depend on hormones but also on increased
concentrations of sucrose.
-
The growth regulators induce the formation
of the new growth regions (sinks), but they are released from the new sinks
& act as strong mobilizing agents.
-
Turgor Sensing in Sugar Transport
-
Strong sinks have high concentrations of solutes
(usually sucrose) in apoplast
-
Strong negative solute potentials result in
diffusion of water out of phloem cells which reducts phloem turgor pressure.
This makes the pressure gradient from source to sink steeper & increases
flow. Reduced turgor in phloem cells at source promotes rapid phloem
loading, which increases rate of transport.
-
Control of Fruit & Vegetable Composition
-
Because developing seeds, fruits, or other
food storage organs transpire at a low rate, they essentially subsist on
phloem sap.
-
Phloem sap composition changes during transport
from leaf to sink .