 We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life.
We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life.
The use of fossils and genetic phylogenies have allowed scientists to reconstruct the history of life and the major epochs and innovations of evolution that have occurred over the last 4 billion years of earth's history. Perhaps the most surprizing thing that we have learned is that we humans are very late arrivals on the evolutionary stage, and that the play had been going on just fine for quite a long while without us. Indeed, the evolution of anything you might recognize as an 'animal' is a fairly recent event in the evolutionary history of the planet; animals have only been present for the last 1/7th of earth history. Although we might think of macroscopic animals and plants as the 'most important' forms of life somehow, it is really the little things - bacteria and archaeans - that keep the biosphere running and allow animals and plants to survive. Without bacteria, you would not digest your food efficiently, plants would have a more difficult time absorbing nutrients, and the nutrients in dead organisms would not be recycled and made available to those of us that are still alive. The basic life processes that drive the biosphere - things like nitrogen fixation, photosynthesis, respiration and energy conversion, and decomposition - are still driven by bacteria. In the first part of this lecture, we'll see that is, indeed, how it has been for the majority of earth history.
We
have summarized some of the major epochs in the history of the Earth before,
but we will review them here and add in important evolutionary events in the
history of life.
4.5 billion years ago - Earth formed (Evidence: meteorites and moon rocks date to this age, dating the condensation of planetissimals in the solar system)
4.0 bya - oldest rocks (The oldest rocks are not as old as the Earth because the Earth's surface was molten - as a result of constant meteorite bombardment and the energy released during the stratification of Earth's core, mantle, and crust. In addition, the Earth is tectonically active, recycling its crust. So, older rocks that may have solidified may have been subducted and melted)
3.5-3.8 bya - origin of life (The oldest fossils, both microfossils and stromatolites, date to 3.5-3.8 bya). These organisms are similar to prokaryotes we see today. However, it is not possible to distinguish whether they are Bacteria or Archaeans. However, from this period forward there are anaerobic and non-oxygenic photosynthetic bacteria.
2.5-2.0 bya - oxygen accumulating ("red bed" sediments form, demonstrating that oxygen gas in the water was oxidizing iron); this is evidence of the evolution of oxygenic photosynthesis. With the accumulation of oxygen, many obligate anaearobes undoubtedly go extinct. However, other bacteria evolve the capacity to USE oxygen as an oxidizing agent, making the harvest of energy from glucose metabolism more efficient... in other words, aerobic respiration evolves.
2.0 bya - eukaryotes evolve through endosymbiotic absorption/tolerance of aerobic bacteria (protomitochondria). With the evolution of eukaryotes comes the evolution of sexual reproduction. These endosymbionts may have been a cellular parasite, like Rickettsia. Subsequent lines of eukaryotes absorbed photosynthetic cyanobacteria, which evolved into symbiotic chloroplasts. The evidence for these theories of eukaryote evolution are the genetic similarity between mitochondria and chloroplasts (that have their own DNA) and various types of bacteria and cyanobacteria. So until this point - representing nearly 1/2 of life's history on this planet - life was exclusively bacterial. The major types of ecological roles, from nitrogen fixation, to photosynthesis, to heterotrophy, to saprophagy (eating dead stuff) all evolved among bacteria - creating food webs and biogeochemical cycles that shunted energy through the biosphere and other earth systems.
0.7 bya - the first animals evolve - sponges and then soft-bodies invertebrate fossils, often radially symmetrical like jellyfish.
0.5 bya - radiation of animals with hard bodies (Cambrian explosion) - beginning of the Paleozoic Era ('old animals')
0.4 bya - evolution of land animals and plants
0.24 bya (240 mya) - Permian extinction; end of the Paleozoic Era
240-65 mya - Mesozoic Era (Age of Dinosaurs - 'middle animals')
65 mya - Cretaceous extinction (meteorite impact off Yucatan Peninsula) - beginning of the 'Cenozoic Era' ('recent animals')
6-5 mya - split of chimp and human lineages
4.5 mya - Australopithecines (1/1000 of earth's history left to present...99.9 % has occurred already).
0.2 mya (200,000) - oldest Homo sapiens fossil. (99.995 % of earth's history has passed).
A. Overview
 These
two domains are ancient; they may even have diverged before 3.5 bya. The problem
with using genetic clocks and genetic similarity to attempt to date this point
of genetic divergence on these groups is that prokaryotes exchange DNA "laterally".
They don't just pass it down a lineage from parental cell to daughter cells;
they can also pass genes from one living bacterium to another through conjugation
or transformation. As such, genetic similarity can occur as a result of lateral
gene transfer between unrelated organisms, and thus the degree of genetic similarity
in particular genes does not necessarily indicate the degree of common ancestry.
For these reasons, it is very difficult to use this particular technique to
date the time of divergence of these two ancient groups.
These
two domains are ancient; they may even have diverged before 3.5 bya. The problem
with using genetic clocks and genetic similarity to attempt to date this point
of genetic divergence on these groups is that prokaryotes exchange DNA "laterally".
They don't just pass it down a lineage from parental cell to daughter cells;
they can also pass genes from one living bacterium to another through conjugation
or transformation. As such, genetic similarity can occur as a result of lateral
gene transfer between unrelated organisms, and thus the degree of genetic similarity
in particular genes does not necessarily indicate the degree of common ancestry.
For these reasons, it is very difficult to use this particular technique to
date the time of divergence of these two ancient groups.
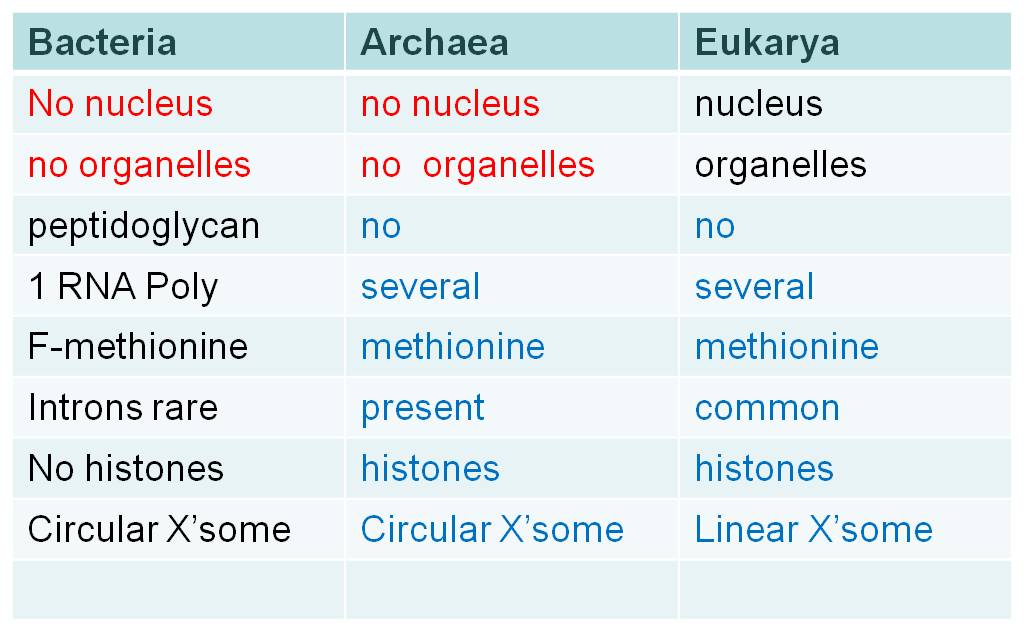
Although they share some fundamental similarities like the lack of a nucleus, the lack of membrane-bound organelles, and circular chromosomes, these two prokaryotic domains differ in some important respects. When they differ, the archaeans are often similar to the Eukarya. For example, bacteria have peptidoglycan in their cell walls, while archaeans (and eukaryotes) do not. Bacteria have only one type of RNA Polymerase; archaeans (and eukaryotes) have several. Bacteria use formyl-methionine as the initiator amino acid in translation, archaeans (and eukaryotes) use methionine. Histones are not associated with bacterial chromosomes, but they are associated with archaean (and eukaryote) chromosomes. Introns are rare in bacterial genes, but are progressively more common in archaeans and eukaryotes. So, there are several fundamental differences between these two groups, and often the archaeans are more similar to eukaryotes than to bacteria.
 B. Metabolic
Diversity in the Prokaryotes
B. Metabolic
Diversity in the Prokaryotes
If we examine the morphological diversity in prokaryotes, we find round (coccus), rod-shaped (bacillus), and spiral (spirochaete). That is not a lot of morphological diversity! So how have the prokaryotes been changing over the last 3.5 by? Well, they have been evolving metabolically. The theme of bacteria diversity is metabolic diversity. There are very few metabolic processes that eukaryotes can perform that prokaryotes cannot. However, the reverse is not true; the prokayotes have a wide range of metabolic diversity - a range that, in many respects, exceeds the metabolic diversity of eukaryotes.
1. Requirements of Oxygen for Cellular Respiration
Consider types of respiration. There
are no anaerobic eukaryotes. Most eukaryotes require oxygen (they have mitochondria,
after all), although some can function without it for a short time (facultative
anaerobes like yeast). Prokaryotes, on the other hand, have a wide range of
'respiratory' strategies:
- some bacteria, like methanogens, cannot live in the presence of oxygen - obligate
anaerobes
- some bacteria can tolerate oxygen
but don't use it - aerotolerant
- some bacteria can live with or without oxygen - facultative aerobes
- some bacteria live only in the presence of oxygen - obilgate aerobes
2. Ecological Roles
- primary producers: bacteria still make a significant contribution to the absorption of CO2 and the production of oxygen on our planet - largely in the oceans. And these oceanic bacteria are the bottom of marine food chains.
- nitrogen-fixing bacteria: can convert N2 into NH3 (ammonia) and then NO3 (nitrates) which can be taken up by plants. Only bacteria can do this; and so all new, natural inputs of nitrogen to the biosphere are by bacteria. (Lots of biologically accessible nitrogen is added to the environment by humans - through fertilizer and the production of NOx compounds by internal combustion engines).
- decomposers: as consumers of dead organic matter, bacteria (and fungi) break down complex organics and harvest energy. And the byproducts of that digestion are mineral nutrients (N, P, K) that are made available for reuptake in the biosphere by plants. So, bacteria are needed for nutrients to cycle through ecosystems.
- pathogenic bacteria: these bacteria get energy from living tissue and destroy cells in the process; they also produce either exotoxins that they secrete, or endotoxins that are bound to the cell wall. Examples of these pathogenic bacterial diseases are: staph infections, salmonella, typhoid, cholera, scarlet fever, tetanus, tuberculosis, botulism, gonorrhea, and syphilis.
- symbiotes: there are gut bacteria in nearly all animals that assist with digestion and may be required for the production and absorption of certain metabolites.There are also bacterial symbionts in plants, protists, and fungi.
As such, bacteria interact in every way possible with other organisms, and form the 'hub' of the biosphere. In some ways, the evolution of life has been an "embellishment" of the bacterial ecosystems that dominated the planet for the first 2.3 billion years of life.
The take home message about prokaryotes is their metabolic diversity. For 1.8 billion years, life on Earth was SOLELY bacterial. How was it evolving? How was it responding to the environment? Well, it seems that life was responding metabolically, adapting by exploiting different environments and energy sources in different ways.
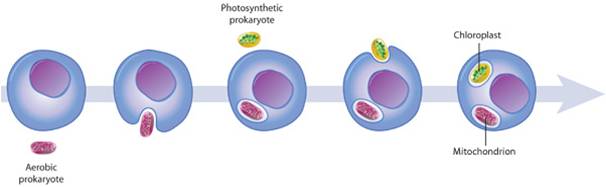
The eukaryotes have been evolving for at least 2.0 billion years. As you might expect (as was argued for the prokaryotes groups), there has been considerable divergence within this group, producing a dramatic range of genetic and metabolic variation. And of course, the multicellular eukaryotes evolved from protistan ancestors - so multicellular eukaryotes (plants, fungi, and animals) are included within this clade. So, the focus will be on the origin of the clade (Eukarya) and the diversity within this clade. We will pay particular attention to the sister groups of the multicellular "kingdoms" that we will consider in greater detail in the remaining lectures. Obviously, the features that distinguish eukaryotes from archaean ancestors are the presence of membrane-bound organelles (mitochondria and chloroplasts) and a nucleus. So, any hypothesis of origin must explain how these defining structures came into existence.
1. Origin of Organelles
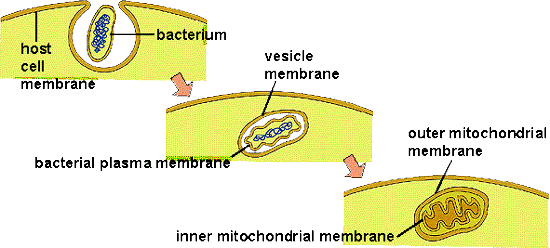
The origin of eukaryotic cells is described by the Endosymbiotic Theory, popularized by Lynn Margulis in the 1960's. It states that organelles are descended from free-living bacterial cells that were absorbed by - or parasitized - archaeans. The evidence for this idea is very strong. Both mitochondria and chloroplasts have double membranes. The outer membranes are similar to the plasma membrane of eukaryotes and archaeans. The inner membrane has certain attributes of bacteria. If a bacterium were absorbed through the process of endocytosis, the bacterium would be surrounded by the membrane of the host's vacuole - creating a double membrane. In addition, both types of organelles have their own DNA, which is circular like bacterial DNA. Both mitochondria and chloroplasts divide by fission, independent of the mitotic division of the eukaryotic cell they inhabit. Over the course of evolutionary history, genes have been exchanged between the organelles and the nucleus, or have been eliminated in the organelle because they also exist in the nucleus. For these reasons, the organelles are functionally dependent on the eukaryotic cell and can no longer live independently for very long.
Additional similarities link these organelles to specific bacteria. Mitochondria not only have their own DNA, but they also produce their own proteins using these DNA recipes. (That should be no surprize!) Protein synthesis occurs on ribosomes within the mitochondria, and their ribosomes are more similar to bacterial ribosomes than to the ribosomes in the cytoplasm of their eukaryotic host cells (produced by the nuclear DNA). Likewise, the genetic code used by mitochondria has a few differences from the universal genetic code; differences also seen in the proteobacteria. Genetic analyses suggest that mitochondria are most closely related to Rickettsia proteobacteria. Rickettsia does not causes "rickets" (a vitamin-D deficiency); rather, Rickettsia bacteria cause a variety of human diseases like many forms of tick-borne fevers (Rocky Mountain spotted fever) and typhus. In addition to having a similar genome to mitochondria, these bacteria are obligate intracellular parasites - so they move into eukaryotic cells and become "organelle-like". In addition, Rickettsia produce ATP in the same way that mitochondria do. So, it is possible that archaeans absorbed bacteria and used them for ATP production, or that the bacteria infected the archaeans to consume their sugars, or some combination of these scenarios. In any case, the bacteria and archaean ultimately benefitted from the interaction, forming a mutualistic or "symbiotic" relationship.
 |
 |
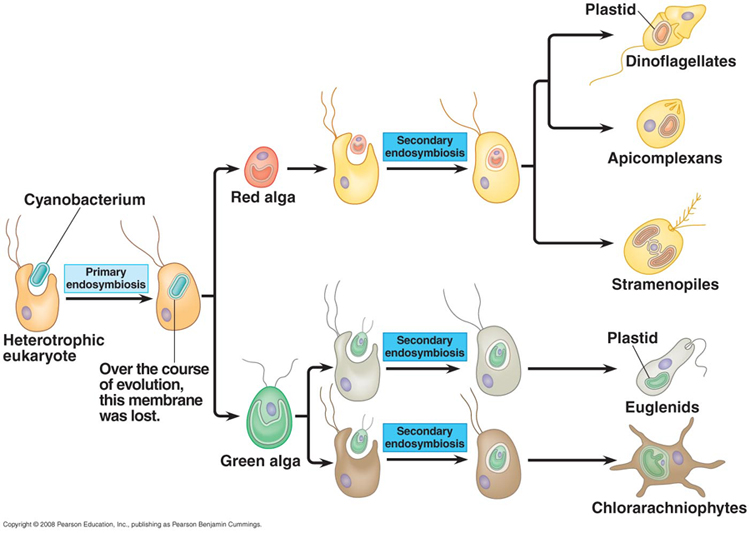
Chloroplasts have an independent, endosymbiotic origin; they are genetically similar to a different group of bacteria called cyanobacteria. Some have a double membrane like mitochondria. However, in some protistan groups, the chloroplasts have 3 or 4 membranes, suggesting multiple rounds of endosymbiosis. So, the current model of chloroplast ancestry suggests a single origin of photosynthetic red and green algae. Green algae were then absorbed (secondary endosymbiosis) by other unrelated protists, creating the euglenids and chlorarachniophytes. These organisms are not related directly through nuclear genetic analysis, but they have very similar, multi-membraned chloroplasts - suggesting independent endocytosis of green algae. A similar process may explain the chloroplast similarity in the stramenopiles, apicomplexans and dinoflagellates - independent endocytosis of a red algae could explain their dissimilar nuclear DNA and their similar, multi-membraned chloroplasts.
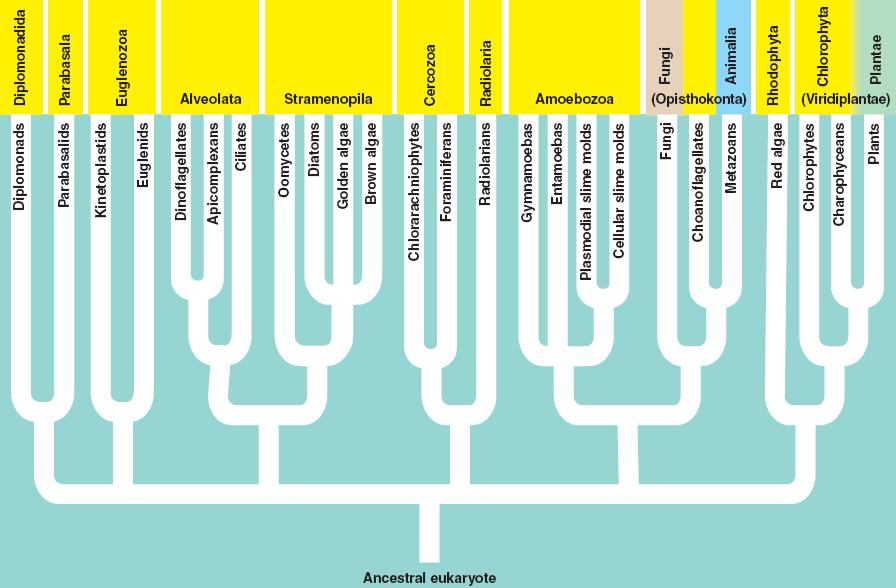
There are several groups of eukarya; most of them contain only single-celled representatives.
 Animal
fossils become much more abundant in the fossil record 550 mya. Although there
are older fossils of soft -bodied jellyfish and worms, the fossil record seems
to 'explode' about 550 mya - an observation that led early Paleotologists to
demarcate this as the beginning of the Paleozoic Era. This explosion probably
occurred because some animals - notably arthropods and molluscs - evolved a
protective body covering (exoskeleton or shell, respectively) that protected
them against the stinging cnidarian predators. The radiation of these groups
left a disproportionate mark on the fossil record because hard parts fossilize
much more readily than soft tissue. So, in part, the 'Cambrian Explosion' is
probably due to sampling bias - organisms evolved that were more likely to fossilize,
so we see more fossils. The evolution of arthropods in the Cambrian led to an
explosion of diversity that is still occurring today - over 85% of the animal
species named to date are Arthropods. In the figure to the right, we see an
artists depiction of the Cambrian Period, with a variety of sponges, cnidarians,
molluscs, and arthropods, including the large predatory arthropod, Anomalocaris - that grasped other animals with its raptorial claws and crushed them in their
circular jaws.
Animal
fossils become much more abundant in the fossil record 550 mya. Although there
are older fossils of soft -bodied jellyfish and worms, the fossil record seems
to 'explode' about 550 mya - an observation that led early Paleotologists to
demarcate this as the beginning of the Paleozoic Era. This explosion probably
occurred because some animals - notably arthropods and molluscs - evolved a
protective body covering (exoskeleton or shell, respectively) that protected
them against the stinging cnidarian predators. The radiation of these groups
left a disproportionate mark on the fossil record because hard parts fossilize
much more readily than soft tissue. So, in part, the 'Cambrian Explosion' is
probably due to sampling bias - organisms evolved that were more likely to fossilize,
so we see more fossils. The evolution of arthropods in the Cambrian led to an
explosion of diversity that is still occurring today - over 85% of the animal
species named to date are Arthropods. In the figure to the right, we see an
artists depiction of the Cambrian Period, with a variety of sponges, cnidarians,
molluscs, and arthropods, including the large predatory arthropod, Anomalocaris - that grasped other animals with its raptorial claws and crushed them in their
circular jaws.
Animals are a fairly recent addition to the living world. They are not very genetically or metabolically diverse when compared to prokaryotes or protists that have much longer evolutionary histories. However, animals are structurally diverse. They have radiated into a wide variety of shapes through changes in developmental patterning. These different shapes have allowed them to exploit a wide range of habitats and resources. Animals are classified into phyla based on their shape or body plan. As heterotrophs, these differences often correlate with differences in feeding structures or locomotory structures that they use to capture food.
From our vantage point as large terrestrial organisms, we have a very skewed view of animal diversity. This is represented by our casual division of animals into 'vertebrates' (like us) and 'invertebrates' (other stuff not like us). These are not phylogenetic groups. The Vertebrata is one sub-phylum of the phylum Chordata, which is one of 32 phyla of animals. The two other sub-phyla of Chordates, and all the other 31 phyla, are 'invertebrate' - and they obviously represent the bulk of animal diversity. Indeed, 95% of all animal species are 'invertebrates', with 85%, as mentioned, occurring in the single phylum Arthropoda. In addition, most animal phyla are exclusively marine - so we rarely encounter them. As you look through the groups pictured at the top of the page (the protostome phyla), you'll see that most animals are rather simple, bilaterally symmetrical, worm-like organisms with different feeding structures.
1. Body Plans
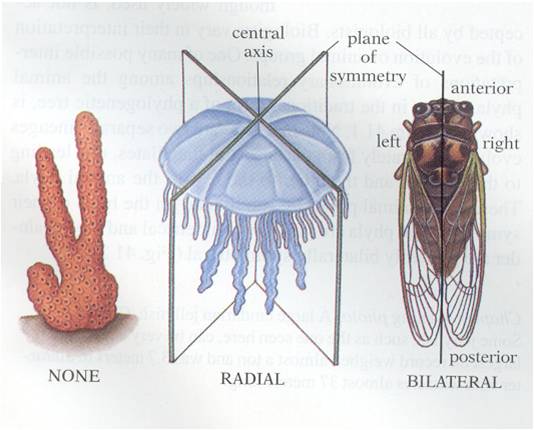 Animals
exibit a wide range of shapes, but these shapes can be classified in three major
groups. First, the sponges are asymmetrical. Second, the cnidarians (jellyfish, corals, sea anemones)
are 'radially symmetrical. This means that, if you are looking down on the organism
from the top, you could cut the organism is half along any axis passing through
the midpoint and the two halves would be be the same. The rest of the animals
are bilaterally symmetrical. This means that there is only one plane of division
that will create to symmetrical 'sides' (bi-lateral = two sides). Given the
fact that the vast majority of animal phyla are bilaterally symmetrical, we
can hypothesize that this evolutionary innovation was functionally important
- allowing animals to work "better", somehow. But how?
Animals
exibit a wide range of shapes, but these shapes can be classified in three major
groups. First, the sponges are asymmetrical. Second, the cnidarians (jellyfish, corals, sea anemones)
are 'radially symmetrical. This means that, if you are looking down on the organism
from the top, you could cut the organism is half along any axis passing through
the midpoint and the two halves would be be the same. The rest of the animals
are bilaterally symmetrical. This means that there is only one plane of division
that will create to symmetrical 'sides' (bi-lateral = two sides). Given the
fact that the vast majority of animal phyla are bilaterally symmetrical, we
can hypothesize that this evolutionary innovation was functionally important
- allowing animals to work "better", somehow. But how?
Bilateral symmetry does more than create two sides; it also means that there is a top and bottom, and a front and back. As heterotrophs, most animals have to moving through the environment in search for their food. Having a 'front' would favor the concentration of sensory systems at this end - so that the animal could perceive the environment that it was about to enter and search for food and avoid predators. So, bilateral symmetry is associated with the formation of a head and the evolution of a brain for processing all this sensory information and directing locomotion. It is no coincidence, therefore, that bilateral symmetry allowed some animals to "get a-head". hahaha.
2. Insects
Half of all species are insects. They are small (so populations split easily), they are tough and can tolerate lots of environments (with an exoskeleton), and they produce lots of eggs so that, if they get somewhere new, they can establish a population quickly. They come in many shapes and sizes, but apparently it's the characteristics listed above that has led to their incredible diversity. They are incrdibly important ecologically. Most are detritivores - they eat dead stuff before bacteria can get it. So, these insects harvest that energy and make it available to larger animals, keeping the energy in the food web. Unfortunately, it seems that insect abundance has declined by 50-70% in the last 40 years, and the loss of insects means the demise of all the other animals that eat them, particularly birds, reptiles, and small mammals.
3. Vertebrates
Vertebrates represent the evolutionary lineage that went in the other direction: not towards a sedentary lifestyle but towards an active lifestyle. For the remainder of this unit, we will look at the evolutionary radiations within the vertebrates. We will see a typical pattern throughout this unit. A new group will evolve a novel trait or way of life. This will allow it to use a new environment, or use the environment in a new way. These lifestyles are called "adaptive zones", and they relate to ecological niches. Creation or entry into a new niche or zone means that there is no competition for resources. Under these conditions, almost any strategy using this new novel innovation will work - typically we see a radiation of new species exploiting this new way of life. As this adaptive zone or niche fills with species, they begin to compete. This leads to 'competitive contraction' of diversity, with only a few winners. The same thing happens in any situation where there are potentially limted resources. A new innovation, like the internet, provides opportunities to new businesses. Many flourish initially - like the many browser providers that were present in the '90's. Anyone remember when Netscape was the primary internet provider? Economists call that period the "dot-com boom", when internet companies could get a start just by colonizing this new open marketplace. As companies grew, they began to compete; and there were winners that maintained their position in the niche, and losers that went extinct. The same thing can happen in ecological systems. Competitive contraction can occur within a clade, or as a consequence of a more efficient, new evolutionary novelty arising. Henry Ford's Model T not only put many other car makers out of business, it also puts LOTS of horse-driven carriage companies out of business! So, as a more advanced, efficient group evolves, we sometimes see contraction in the abundance of ancestral forms... just as Darwin predicted.
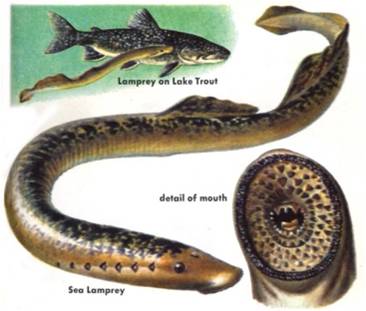 The
jawless fishes: The vertebrates are distinguished by having an internal supportive spine that
is subdivided into units called vertebrae. This segmentation of the vertebral
column allows the animal to bend and move with more precision; with some parts
of the body moving while other parts remain stationary. The most primitive members
of this clade are the jawless fishes. The modern representatives are the hagfish
and the lampreys. The hagfish actually lack a subdivided vertebral column, but
they do have a skull surrounding their brain. As such, sometimes the "vertebrata"
are more specifically described as having a skull (to include the hagfish).
Both hagfish and lampreys have immature stages that look almost identical to
cephalochordates, also suggesting their primitive position within this clade.
So, the jawless fish evolve in the Cambrian and radiate in the Ordovician and
Silurian Periods, with many large, detritivorous and filter feeding species
scouring the benthos for food. In more advanced forms, the mouth cavity and
pharynx evolved for feeding, relieving the gills of this filtering function.
These jawless fish probably sucked in small invertebrates by rapidly opening
their mouth.
The
jawless fishes: The vertebrates are distinguished by having an internal supportive spine that
is subdivided into units called vertebrae. This segmentation of the vertebral
column allows the animal to bend and move with more precision; with some parts
of the body moving while other parts remain stationary. The most primitive members
of this clade are the jawless fishes. The modern representatives are the hagfish
and the lampreys. The hagfish actually lack a subdivided vertebral column, but
they do have a skull surrounding their brain. As such, sometimes the "vertebrata"
are more specifically described as having a skull (to include the hagfish).
Both hagfish and lampreys have immature stages that look almost identical to
cephalochordates, also suggesting their primitive position within this clade.
So, the jawless fish evolve in the Cambrian and radiate in the Ordovician and
Silurian Periods, with many large, detritivorous and filter feeding species
scouring the benthos for food. In more advanced forms, the mouth cavity and
pharynx evolved for feeding, relieving the gills of this filtering function.
These jawless fish probably sucked in small invertebrates by rapidly opening
their mouth.
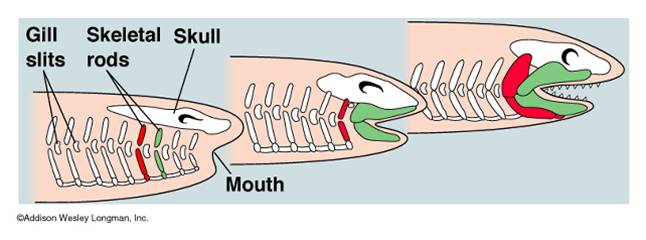 The
Gnathostomes: The Jawed Vertebrates: Jawed fishes evolved
in the Silurian and came to dominate and radiate in the Devonian - the "Age
of Fishes". Jaws evolved from the anterior gill arches, which flex and
became associated withe the mouth. Jaws allowed fish to become more efficient
predators, killing things bigger than could fit in their mouth through suction,
or strong enough to escape the suctioning force. In short, jaws were adaptive
because fish now became mobile, active predators - entering a niche previously
only occupied by large invertebrate predators. The first jawed fish radiation
were the placoderms, symbolized by the frightening Arthrodire predators like Dunkleosteus, with its huge slicing jaw plates. The placoderms were
very heavy, however, and were displaced by more efficient swimmers - the cartilaginous
jawed fishes (Class Chondrichthyes). Modern representatives of this group are
sharks and rays.
The
Gnathostomes: The Jawed Vertebrates: Jawed fishes evolved
in the Silurian and came to dominate and radiate in the Devonian - the "Age
of Fishes". Jaws evolved from the anterior gill arches, which flex and
became associated withe the mouth. Jaws allowed fish to become more efficient
predators, killing things bigger than could fit in their mouth through suction,
or strong enough to escape the suctioning force. In short, jaws were adaptive
because fish now became mobile, active predators - entering a niche previously
only occupied by large invertebrate predators. The first jawed fish radiation
were the placoderms, symbolized by the frightening Arthrodire predators like Dunkleosteus, with its huge slicing jaw plates. The placoderms were
very heavy, however, and were displaced by more efficient swimmers - the cartilaginous
jawed fishes (Class Chondrichthyes). Modern representatives of this group are
sharks and rays.
 The
Devonian also saw the radiation of the most successful group of fish on earth,
the bony fish (Class Osteichthyes). Unlike cartilage, bone is hollow and light;
it is also more rigid than cartilage, providing more resistance and efficiency
to muscles. The bony fish also had a swim bladder, whcih they used to maintain
neutral bouyancy in the water column. This means that they can maintain position
without swimming; Condrichthyes must swim to maintain vertical position in the
water column - otherwise they sink. These two adaptations of the bony fishes
made them more energetically efficient and faster than the cartilaginous fish,
and the bony fishes have radiated to the point where they represent 40% of all
vertebrate species today (not just fish - vertebrates..!!). Two important clades
evolved within the bony fishes: 1) the ray-finned fishes that came to dominate
the aquatic environments of the planet; and 2) the lob-finned fishes that radiated
into shallow-water environments. The lobe-finned fishes, swimming in shallow,
oxygen-poor waters of the Devonian, gulping air at the surface and pulling themselves
along on their forefins, evolved into the terrestrial tetrapods. Arthropods
had already colonized land, so there was ample food and oxygen for a large terrestrial
predator. As you have seen, the fossil record nicely documents the transition
from fish to these primitive amphibians.
The
Devonian also saw the radiation of the most successful group of fish on earth,
the bony fish (Class Osteichthyes). Unlike cartilage, bone is hollow and light;
it is also more rigid than cartilage, providing more resistance and efficiency
to muscles. The bony fish also had a swim bladder, whcih they used to maintain
neutral bouyancy in the water column. This means that they can maintain position
without swimming; Condrichthyes must swim to maintain vertical position in the
water column - otherwise they sink. These two adaptations of the bony fishes
made them more energetically efficient and faster than the cartilaginous fish,
and the bony fishes have radiated to the point where they represent 40% of all
vertebrate species today (not just fish - vertebrates..!!). Two important clades
evolved within the bony fishes: 1) the ray-finned fishes that came to dominate
the aquatic environments of the planet; and 2) the lob-finned fishes that radiated
into shallow-water environments. The lobe-finned fishes, swimming in shallow,
oxygen-poor waters of the Devonian, gulping air at the surface and pulling themselves
along on their forefins, evolved into the terrestrial tetrapods. Arthropods
had already colonized land, so there was ample food and oxygen for a large terrestrial
predator. As you have seen, the fossil record nicely documents the transition
from fish to these primitive amphibians.
 The
Tetrapods: The
tetrapod clade includes all vertebrates descended from the first animals to
walk on land. The radiation of early tetrapods is very impressive; the adaptive
zone of the Carboniferous swamps was very large and well-suited to amphibian
animals reliant on water to lay their eggs. In addition, the huge amount of
biological productivity (lots of photosynthesis and less decompostion, remember?)
led to large populations of herbivorous insects that were eay prey for large
land vertebrates. The Carboniferous could well be called the "Age of Amphibians".
Many groups of early 'amphibians' radiate during this period, collectively called
the 'stem tetrapods'. However, the dry periods of the Permian and Mesozoic were
not optimal for these animals limited to areas near water. Today, three groups
of amphibians remain. The Caecilians are unusual, rare, legless amphibians.
Salamanders are the second group, and the giant Japanese salamander is the largest
living amphibian - reaching six feet in length! The most diverse group is the
Anura - mening "tailless" - the frogs and toads. Most amphibians have
gills or lungs at some point in their life cycle, but many also (or exclusively)
respire through their skin. For this to happen, the gases must diffuse into
a thin film of water, and then move across cell membranes by diffusion. So,
most amphibians need to keep their skin moist to respire; and it also means
that their skin, as an absorptive surface, is also prone to absorb toxins in
the environment. For these reasons, amphibians are particularly sensitive to
pollutants and environmental contaminants. Over the last 50 years, there has
been a dramatic decline in amphibian populations worldwide, probably due to
exposure to human-produced pollution, climate change, and chytrid fungal infections.
It is tough to say which of these variables is 'most' responsible for the decline.
In all likelihood, each factor increases sensitivity to the other two. In any
case, these three factors are having a decimating effect on amphibian populations.
The
Tetrapods: The
tetrapod clade includes all vertebrates descended from the first animals to
walk on land. The radiation of early tetrapods is very impressive; the adaptive
zone of the Carboniferous swamps was very large and well-suited to amphibian
animals reliant on water to lay their eggs. In addition, the huge amount of
biological productivity (lots of photosynthesis and less decompostion, remember?)
led to large populations of herbivorous insects that were eay prey for large
land vertebrates. The Carboniferous could well be called the "Age of Amphibians".
Many groups of early 'amphibians' radiate during this period, collectively called
the 'stem tetrapods'. However, the dry periods of the Permian and Mesozoic were
not optimal for these animals limited to areas near water. Today, three groups
of amphibians remain. The Caecilians are unusual, rare, legless amphibians.
Salamanders are the second group, and the giant Japanese salamander is the largest
living amphibian - reaching six feet in length! The most diverse group is the
Anura - mening "tailless" - the frogs and toads. Most amphibians have
gills or lungs at some point in their life cycle, but many also (or exclusively)
respire through their skin. For this to happen, the gases must diffuse into
a thin film of water, and then move across cell membranes by diffusion. So,
most amphibians need to keep their skin moist to respire; and it also means
that their skin, as an absorptive surface, is also prone to absorb toxins in
the environment. For these reasons, amphibians are particularly sensitive to
pollutants and environmental contaminants. Over the last 50 years, there has
been a dramatic decline in amphibian populations worldwide, probably due to
exposure to human-produced pollution, climate change, and chytrid fungal infections.
It is tough to say which of these variables is 'most' responsible for the decline.
In all likelihood, each factor increases sensitivity to the other two. In any
case, these three factors are having a decimating effect on amphibian populations.
The Amniota: Within the tetrapods, a new type of animal evolved in the Carboniferous; an animal that laid an egg surrounded by a series of membranes and a shell. These layers served to protect the developing embryo and yolk sac from dessication, allowing these species to colonize land further from water and exploit food resources and habitats that amphibians could not reach. This clade of animals is the amniotes, and their evolutionary innovation is the amniotic egg. The primitive condition within this group probably produced a leathery covering, much like turtles. A more rigid, desiccation resistant egg like birds have is probably a derived trait, as is the retention of the egg and live bearing of young, as seen in some snakes, and most mammals. The amniotes radiated during the Permian Period, when the drying of the unified land mass of Pangaea gave them a decided advantage over the ancestral, amphibious tetrapods. Three groups evolved at this time, distinguished by their skull morphology.
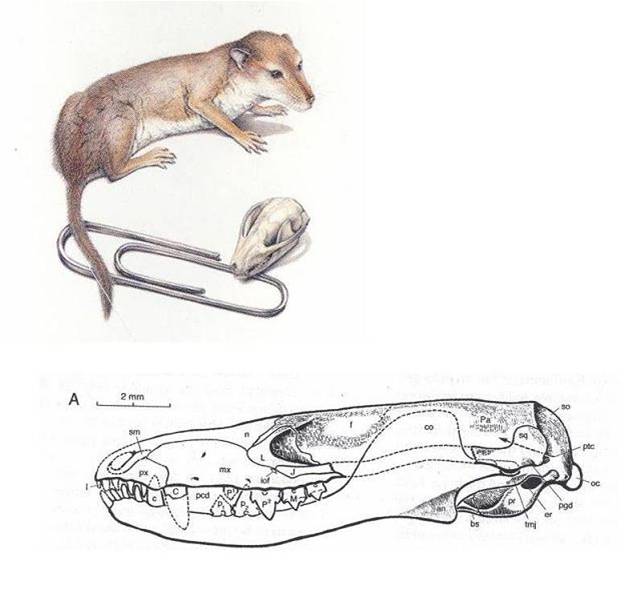 The
Synapsids and the Mammals: The first group to dominate
were the synapsids. They dominated during the Permian, and were represented
by the great "sail-finned" Pelycosaurs and the Therapsid lineages
(the Gorgonopsians, the Dicynodonts, and the Cynodonts). The synapsid lineage
that is alive today are the Mammals. The transition from ancestral to derive
synapsids is well preserved in the fossil record, documenting the evolution
of the inner ear from the ancestral jaw bones, and the evolution of complex
and specialized dentition. The first true mammals are the Morganucodonts that
evolved in the Jurassic Period of the Mesozoic, about 200 mya. Ancestral synapsids
were large carnivores that dominated the landscape; a niche that would be filled
by the diapsid dinosaurs during the Mesozoic. The mammals were small, noctural
insectivores and scavengers. They radiated into these niches in the Jurassic,
perhaps because their endothermy allowed them to use the cool night more effectively
and under cover of darkness from larger predators. The surviving lineage of
this radiation are the three species of monotremes alive today: the platypus
and two species of echidna. These mammals lay eggs. Their offspring hatch, and
then nuzzle the belly of their mothers. Their mothers have sweat glands that
have evolved to produce a nutrient rich milk - they are mammary glands. However,
even the modern representatives lack nipples on these glands - so they "sweat"
milk onto their belly and the the hatchlings lick it up. Placental and marsupial
mammals evolved later, in the Cretaceous Period. Marsupials do not lay eggs;
they give live birth to a very small, embryonic newborn. The newborn climbs
from the genital opening to the pouch - a flap of skin covering the nipples
of the mammary glands. The embryo attaches to a nipple and feeds nearly continuously,
completing development in the pouch. This allows the mother to carry the embryo
with her for a longer period; there are no eggs that must be left periodically
for the mother to feed. The last group of mammals, the placentals, also retain
the embryo and give live birth. However, they retain the embryo much longer,
often until the offspring is capable of independent activity. The efficient
feeding of the embryo is facilitated by the placenta - which allows nutrients
in the bloodstream of the mother to pass directly into the bloodstream of the
offspring. This is far more efficient, energetically, than the mother turning
the nutrients into milk, that must be consumed and digested by the offspring.
So, placental mammals can develop more rapidly than marsupials. Some placentals
produce offspring that can walk immediately; these are precocial young. Other
species, like cats and humans, produce offspring that still need considerable
parental care. These offspring are called altricial.
The
Synapsids and the Mammals: The first group to dominate
were the synapsids. They dominated during the Permian, and were represented
by the great "sail-finned" Pelycosaurs and the Therapsid lineages
(the Gorgonopsians, the Dicynodonts, and the Cynodonts). The synapsid lineage
that is alive today are the Mammals. The transition from ancestral to derive
synapsids is well preserved in the fossil record, documenting the evolution
of the inner ear from the ancestral jaw bones, and the evolution of complex
and specialized dentition. The first true mammals are the Morganucodonts that
evolved in the Jurassic Period of the Mesozoic, about 200 mya. Ancestral synapsids
were large carnivores that dominated the landscape; a niche that would be filled
by the diapsid dinosaurs during the Mesozoic. The mammals were small, noctural
insectivores and scavengers. They radiated into these niches in the Jurassic,
perhaps because their endothermy allowed them to use the cool night more effectively
and under cover of darkness from larger predators. The surviving lineage of
this radiation are the three species of monotremes alive today: the platypus
and two species of echidna. These mammals lay eggs. Their offspring hatch, and
then nuzzle the belly of their mothers. Their mothers have sweat glands that
have evolved to produce a nutrient rich milk - they are mammary glands. However,
even the modern representatives lack nipples on these glands - so they "sweat"
milk onto their belly and the the hatchlings lick it up. Placental and marsupial
mammals evolved later, in the Cretaceous Period. Marsupials do not lay eggs;
they give live birth to a very small, embryonic newborn. The newborn climbs
from the genital opening to the pouch - a flap of skin covering the nipples
of the mammary glands. The embryo attaches to a nipple and feeds nearly continuously,
completing development in the pouch. This allows the mother to carry the embryo
with her for a longer period; there are no eggs that must be left periodically
for the mother to feed. The last group of mammals, the placentals, also retain
the embryo and give live birth. However, they retain the embryo much longer,
often until the offspring is capable of independent activity. The efficient
feeding of the embryo is facilitated by the placenta - which allows nutrients
in the bloodstream of the mother to pass directly into the bloodstream of the
offspring. This is far more efficient, energetically, than the mother turning
the nutrients into milk, that must be consumed and digested by the offspring.
So, placental mammals can develop more rapidly than marsupials. Some placentals
produce offspring that can walk immediately; these are precocial young. Other
species, like cats and humans, produce offspring that still need considerable
parental care. These offspring are called altricial.
The Diapsids: The other major group of amniotes are the diapsids. They radiated into two major groups, the lepidosaurs that walked with their legs splayed out and the archosaurs that walked with their legs underneath.
 The
archosaurs would dominate during the Mesozoic Era, with the great
radations of the Crocodylomorphs, Pterosaurs, and Dinosaurs. Two groups of Archosaurs
persists today - the crocodilians (caimen, gavials, alligators, and crocodiles),
and the birds. Although several groups of dinosaurs evolved feathers - probably
first for attracting mates or communicating, and then for insulation - only
in the birds did these feathers evolve for powered flight. Associated shared
derived characters are light, hollow, fused bones, the fusion of digits in the
forelimbss, a keeled sternum for anchoring large breast muscles used for flight,
and a 'wishbone' - a union of the two clavicles that acts like a spring during
the flight stroke. The loss of teeth is also a derived trait in this group,
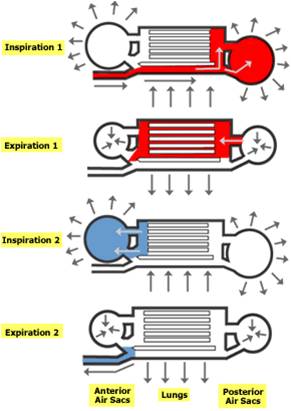
as is the evolution of an interesting respiratory system of air sacs. Birds
have a one way lung. Air that's inhaled goes to a posterior air sac. On the
first exhalation, air is moved forward through the lung. The second inhalation
refills the posterior air sac, and the next exhalation pulls deoxygenated air
forward into the anterior air sac and out of the animal. This modification,
like a one-way digestive tract, improves the efficiency of the organ. Now, the
air in the lung is always oxygenated; it is not mixed with deoxygenated air
like in the sac-like lungs of other animals. It's not surprizing that birds
would evolve this system, as the metabolic demands of flight are much more significant
than other forms of locomotion.
The
archosaurs would dominate during the Mesozoic Era, with the great
radations of the Crocodylomorphs, Pterosaurs, and Dinosaurs. Two groups of Archosaurs
persists today - the crocodilians (caimen, gavials, alligators, and crocodiles),
and the birds. Although several groups of dinosaurs evolved feathers - probably
first for attracting mates or communicating, and then for insulation - only
in the birds did these feathers evolve for powered flight. Associated shared
derived characters are light, hollow, fused bones, the fusion of digits in the
forelimbss, a keeled sternum for anchoring large breast muscles used for flight,
and a 'wishbone' - a union of the two clavicles that acts like a spring during
the flight stroke. The loss of teeth is also a derived trait in this group,
as is the evolution of an interesting respiratory system of air sacs. Birds
have a one way lung. Air that's inhaled goes to a posterior air sac. On the
first exhalation, air is moved forward through the lung. The second inhalation
refills the posterior air sac, and the next exhalation pulls deoxygenated air
forward into the anterior air sac and out of the animal. This modification,
like a one-way digestive tract, improves the efficiency of the organ. Now, the
air in the lung is always oxygenated; it is not mixed with deoxygenated air
like in the sac-like lungs of other animals. It's not surprizing that birds
would evolve this system, as the metabolic demands of flight are much more significant
than other forms of locomotion.
The Lepidosaurs also radiated during the Mesozoic, but did not do so as dramatically as the Archosaurs. Today, the surviving representatives are the Squamata (lizards and the derived snakes) and their sister group, the Sphenodonts with two surviving relict species of Tuatara native to New Zealand. The tuatara maintains the most primitive amniote characteristics, and so is very valuable in studying the phylogeny of diapsid reptiles. The derived squamates, the modern lizards and snakes, have scales, a double hinged jaw (particularly important in snakes), and a hinge on the top of their skull.
That concludes a look at the phyla of animals. Next we will take a brief look at the evolutionary history of our own small twig on the mammalian limb.
Study Questions:
1. Known the timeline of major events in Earth history.
2. bacteria and archaea are more genetically diverse than eukaryotes. Why? And what does this diversity code for?
3. All eukaryotes are aerobic. List four types of metabolism that prokaryotes have with respect to oxygen.
4. Describe five ecological roles that prokaryotes play in the environment.
5. Describe the endosymbiotic theory of organelle origin, and provide evidence for this theory.
4. Why is bilateral symmetry, rather than radial symmetry, associated with "cephalization" in motile organisms?
5. Why are insects so diverse?
6. How did the jaw evolve and what benefit did it provide?
7. What were the probable benefits of colonizing land?
8. What is an amniotic egg and what benefit does it provide? Why did amniotes radiate in the Permian?
9. Describe the reproductive differences among the monotremes, marsupials, and placentals. What advantages were there for marsupial and then placental reproductive strategies?
10. Explain how a bird's respiratory system is more efficient than a mammal's.
11. What challenges face an aquatic algae as it tries to acquire light, gases, and nutrients?