 The
Diversity of Life The
Diversity of Life
The
Diversity of Life The
Diversity of Life 5. Major Phyla
a. Phylum Porifera: Sponges
b. Phylum Cnidaria: Hydra, anemones, corals, jellyfish
The "Bilateria"
The remaining animal phyla are bilaterally symmetrical organisms. Their symmetry is encoded by Hox genes that establish the anterior-posterior axis of the body. As mentioned previously, bilateral symmetry is advantageous because it establishes a polarity to the animal - and sense systems and neural integration can be concentrated at the anterior end.
The bilaterally symmetrical animals have three true tissue layers (triploblastic), adding mesoderm to endo- and ectoderm. They form two major clades, distinguished by whether the blastopore developes into the mouth (Protostomes) or the anus (Deuterostomes) in the adult. The protostomes are further divided by other developmental characteristics. One major clade, the Ecdysozoa, must shed a hard cuticle or exoskeleton in order to grow. The other clade, the Lophotrochozoans, is distinguished by having either a rake-like feeding apparatus ('lophophore' - seen in some of the pictures in the lower two rows, above), or a specific larval stage known as a 'trochophore'. As you can see from the phylogeny above, there are many groups whose phylogenic relationships are unknown at this time.
Protostomes: Lophotrochozoans
c. Phylum Platyhelminthes: Flatworms
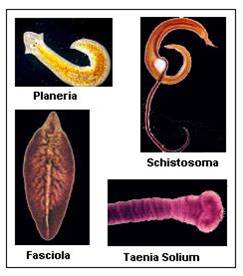 The
flatworms are a rather primitive group of bilaterally symmetrical organisms
that maintain some of the basic characteristics of cnidarians. Like cnidarians,
they have a digestive cavity with only one opening; they do not have a one-way
digestive tract. However, the gut of flatworms is highly bifurcated and convoluted,
with pockets that radiate out into the deep tissue on the sides of the animal.
This is probably adaptive in this organism because it insures that no cell is
too far from the gut (and digested nutrients). Flatworms also have ameobocytes
like sponges and cnidarians, too; these transport nutrients between the endodermal
cells lining the gut and the rest of the tissues in the animal. Digestion is
both intracellular and extracellular, as in cnidarians. The major evolutionary
innovations in this group are associated with bilaterality. The nervous system
and sensory organs (eyespots and chemoreceptive auricles) are concentrated towards
the anterior end of the animal, creating a head (of sorts). Flatworms are a
diverse group, with free-living and parasitic forms. Planaria live in marine,
freshwater, and terrestrial environments. Flukes are parasitic, like the liver
flukes and blood flukes (Schistosomes) we saw in lab. Most of these species
have at least one intermediate host and a complex life cycle. At some point
in the life cycle there is usually an asexual stage in which thousands of offspring
can be produced asexually. This is particularly important for parasites that
infect different hosts in sequence, because the probability of finding new hosts
is low. Tapeworms are another group of parastic flatworms. Over 1000 species
have been described, and every vertebrate species is infected by at least one
species of tapeworm. Humans are infected by three, with sheep, pigs, cattle,
and fish acting as intermediate hosts. Tapeworms are weird. Humans ingest living
cysts in undercooked beef, lamb, pork, and fish. The immature worm hatches from
the cysts and attached to the wall of the intestine with the hooks on the scolex
- the head segment. As the worm matures and grows, it produces body segments
called proglottids. Each proglottid contains male and female reproductive organs.
As the worm creates new proglottids behind the head, older proglottids that
are farther from the head mature. The worm can be over 12m long in humans; some
grow to over 30m in length! Each proglottid is metabolically independent, absorbing
nutrients by diffusion, directly from the hosts intestinal tract. The high SA/V
ratio of these flatworms aids in this parasitic lifestyle. Each proglottid produces
egg and sperm; sperm are received by proglottids and the development of fertilized
eggs occurs in each proglottid. Eventually, the mature proglottids are shed
by the worm, and are shed from the host in feces. The proglottids are eaten
by the secondary host (sheep, pigs, cattle, and fish). The larvae hatch, bore
through the intestinal wall, and migrate to muscle tissue where they encyst.
Consumption of undercooked meat by another person completes the life cycle.
The
flatworms are a rather primitive group of bilaterally symmetrical organisms
that maintain some of the basic characteristics of cnidarians. Like cnidarians,
they have a digestive cavity with only one opening; they do not have a one-way
digestive tract. However, the gut of flatworms is highly bifurcated and convoluted,
with pockets that radiate out into the deep tissue on the sides of the animal.
This is probably adaptive in this organism because it insures that no cell is
too far from the gut (and digested nutrients). Flatworms also have ameobocytes
like sponges and cnidarians, too; these transport nutrients between the endodermal
cells lining the gut and the rest of the tissues in the animal. Digestion is
both intracellular and extracellular, as in cnidarians. The major evolutionary
innovations in this group are associated with bilaterality. The nervous system
and sensory organs (eyespots and chemoreceptive auricles) are concentrated towards
the anterior end of the animal, creating a head (of sorts). Flatworms are a
diverse group, with free-living and parasitic forms. Planaria live in marine,
freshwater, and terrestrial environments. Flukes are parasitic, like the liver
flukes and blood flukes (Schistosomes) we saw in lab. Most of these species
have at least one intermediate host and a complex life cycle. At some point
in the life cycle there is usually an asexual stage in which thousands of offspring
can be produced asexually. This is particularly important for parasites that
infect different hosts in sequence, because the probability of finding new hosts
is low. Tapeworms are another group of parastic flatworms. Over 1000 species
have been described, and every vertebrate species is infected by at least one
species of tapeworm. Humans are infected by three, with sheep, pigs, cattle,
and fish acting as intermediate hosts. Tapeworms are weird. Humans ingest living
cysts in undercooked beef, lamb, pork, and fish. The immature worm hatches from
the cysts and attached to the wall of the intestine with the hooks on the scolex
- the head segment. As the worm matures and grows, it produces body segments
called proglottids. Each proglottid contains male and female reproductive organs.
As the worm creates new proglottids behind the head, older proglottids that
are farther from the head mature. The worm can be over 12m long in humans; some
grow to over 30m in length! Each proglottid is metabolically independent, absorbing
nutrients by diffusion, directly from the hosts intestinal tract. The high SA/V
ratio of these flatworms aids in this parasitic lifestyle. Each proglottid produces
egg and sperm; sperm are received by proglottids and the development of fertilized
eggs occurs in each proglottid. Eventually, the mature proglottids are shed
by the worm, and are shed from the host in feces. The proglottids are eaten
by the secondary host (sheep, pigs, cattle, and fish). The larvae hatch, bore
through the intestinal wall, and migrate to muscle tissue where they encyst.
Consumption of undercooked meat by another person completes the life cycle.
d. Phylum Annelida: Segmented worms
 Annelids
include three major groups - the polychaete worms (like we used in lab as the
outgroup for assessing shared derived traits in arthropods), earthworms (like
we dissected in lab), and leeches. Annelids inhabit marine, freshwater, and
terrestrial environments. The annelids are an ancient group, but their soft
bodies do not fossilize well. However, there are unambiguous annelids that date
to the Cambrian, and the putative polychaete Dicksonia dates from the
Vendian Period 600 mya. With the Annelida we see a fundamental change in body
plan - segmentation. Like gene duplication, the replication of body units into
separate segments easily allows for body region to specialize, or to evolve
a new function in one area without compromising the ability to perform original
functions in other segments. This redundancy is easily seen in the anterior
region of earthworms, where each segment contains a heart. Likewise, each segment
throughout the animal contains its own excretory units - nephridia. Each segment
of polychaetes has parapodia - 'side-feet' used for locomotion and respiration
(increasing the surface area for gas exchange). Annelids have a true body cavity,
they are not solid cells from the gut to the outer wall. This cavity provides
room for outpocketings of the digestive tract and other organs. In addition,
this fluid-
Annelids
include three major groups - the polychaete worms (like we used in lab as the
outgroup for assessing shared derived traits in arthropods), earthworms (like
we dissected in lab), and leeches. Annelids inhabit marine, freshwater, and
terrestrial environments. The annelids are an ancient group, but their soft
bodies do not fossilize well. However, there are unambiguous annelids that date
to the Cambrian, and the putative polychaete Dicksonia dates from the
Vendian Period 600 mya. With the Annelida we see a fundamental change in body
plan - segmentation. Like gene duplication, the replication of body units into
separate segments easily allows for body region to specialize, or to evolve
a new function in one area without compromising the ability to perform original
functions in other segments. This redundancy is easily seen in the anterior
region of earthworms, where each segment contains a heart. Likewise, each segment
throughout the animal contains its own excretory units - nephridia. Each segment
of polychaetes has parapodia - 'side-feet' used for locomotion and respiration
(increasing the surface area for gas exchange). Annelids have a true body cavity,
they are not solid cells from the gut to the outer wall. This cavity provides
room for outpocketings of the digestive tract and other organs. In addition,
this fluid-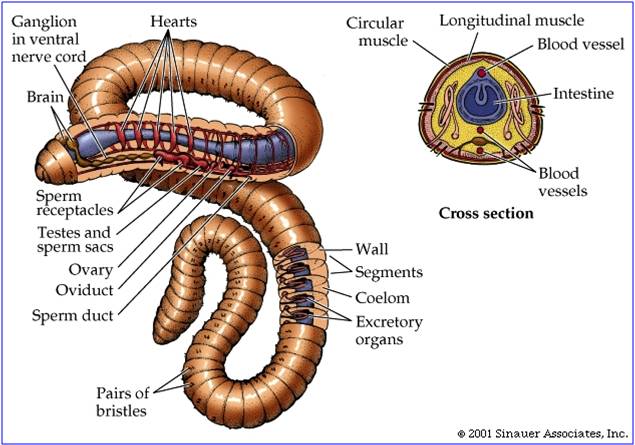 filled
space can act as a hydrostatic skeleton that allows for refined, peristaltic
locomotion. Water is incompressible, so muscles contracting in one part of the
body push the water in the coelom somewhere else, like towards the front of
the animal which extends the body forward. Annelids also have another important
difference with the other phyla we have considered - they have a digestive tract
rather than a gut cavity. In a gut cavity, new food is added and is mixed with
digesting food; there is no separation and so there can be no specialization
of digestive function. In a tract, where food enters at one end of the animal
(mouth) and passes in one direction towards the exit (anus), there can be specialization
of function along the route - creating a "dis-assembly line" that
breaks food down and harvests energy more efficiently. We see this specialization
in earthworms, where the mucular pharynx contracts to extend the mouth for ingestion,
the crop that stores food and delivers it evenly to the gizzard, which grinds
the food into smaller particles (increasing its surface area for the action
of enzymes), and the intestine where digestion and the absorption of nutrients
occurs. Earthworms eat soil, digesting the fungi, bacteria, organic molecules
and protists. They pass fecal pellets that contain the mineral components and
unabsorbed organics. By creating these pellets, they improve the ability of
water and air to travel through the soil, increasing the uptake of water and
soluble nutrients, and increasing gas exchange by plant roots. This increases
the growth rate of plants and increases the productivity of the community.
filled
space can act as a hydrostatic skeleton that allows for refined, peristaltic
locomotion. Water is incompressible, so muscles contracting in one part of the
body push the water in the coelom somewhere else, like towards the front of
the animal which extends the body forward. Annelids also have another important
difference with the other phyla we have considered - they have a digestive tract
rather than a gut cavity. In a gut cavity, new food is added and is mixed with
digesting food; there is no separation and so there can be no specialization
of digestive function. In a tract, where food enters at one end of the animal
(mouth) and passes in one direction towards the exit (anus), there can be specialization
of function along the route - creating a "dis-assembly line" that
breaks food down and harvests energy more efficiently. We see this specialization
in earthworms, where the mucular pharynx contracts to extend the mouth for ingestion,
the crop that stores food and delivers it evenly to the gizzard, which grinds
the food into smaller particles (increasing its surface area for the action
of enzymes), and the intestine where digestion and the absorption of nutrients
occurs. Earthworms eat soil, digesting the fungi, bacteria, organic molecules
and protists. They pass fecal pellets that contain the mineral components and
unabsorbed organics. By creating these pellets, they improve the ability of
water and air to travel through the soil, increasing the uptake of water and
soluble nutrients, and increasing gas exchange by plant roots. This increases
the growth rate of plants and increases the productivity of the community.
e. Phylum Mollusca: Chitons, Bivalves, Snails, and Cephalopods
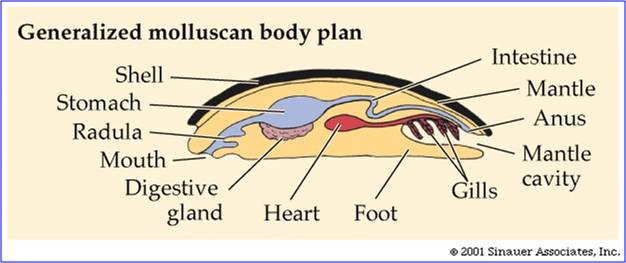 The
molluscs are also an ancient group, dating to the late pre-Cambrian Period (about
560 mya). Shelled molluscs, protected from cnidarian predators, radiate in the
Cambrian. The molluscan body plan is fairly simple, best seen in the most primitive
extant lineage, the Polyplacophorans (Chitons). They are bilaterally symmetrical
with a complete digestive tract. Their body is covered by a hood-like structure
called the mantle. This mantle creates a pocket between the body of the animal
and the hood. Gills are present in this mantle cavity. The bottom of the animal
is a muscular foot. In many molluscs, the mantle secretes a shell. In the polyplacophorans,
the shell is composed of 8 units or plates, suggesting the segmented ancestry
of the group. However, more derived molluscs have lost most indications of segmentation.
Although molluscs take a wide variety of shapes, from snails to clams to squid,
all of these shapes are just variations on this basic theme.
The
molluscs are also an ancient group, dating to the late pre-Cambrian Period (about
560 mya). Shelled molluscs, protected from cnidarian predators, radiate in the
Cambrian. The molluscan body plan is fairly simple, best seen in the most primitive
extant lineage, the Polyplacophorans (Chitons). They are bilaterally symmetrical
with a complete digestive tract. Their body is covered by a hood-like structure
called the mantle. This mantle creates a pocket between the body of the animal
and the hood. Gills are present in this mantle cavity. The bottom of the animal
is a muscular foot. In many molluscs, the mantle secretes a shell. In the polyplacophorans,
the shell is composed of 8 units or plates, suggesting the segmented ancestry
of the group. However, more derived molluscs have lost most indications of segmentation.
Although molluscs take a wide variety of shapes, from snails to clams to squid,
all of these shapes are just variations on this basic theme.
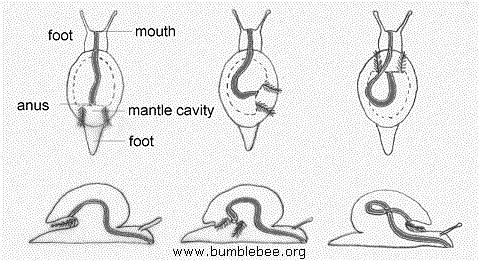 Snails
have put a 'twist' on molluscan evolution. The shell and mantle rotate during
development, placing the mantle cavity at the back of the animal. Many snails
are grazers; they scrape algae off rocks with a file-like tongue called a radula.There
are many predatory snails, however. Oyster drills have a tough radula, and they
scrape a hole through oyster shells. They then extend their proboscis through
the hole and consume the oyster. Coneshells, in the genus Conus, have
a modified radula that takes the shape and function of a harpoon. They stab
fish and inject a potent toxin; some Conus snail toxins are also lethal
to humans. Almost all molluscs are marine or aquatic; only some snails have
colonized land.The most coloful molluscs are nudibranchs (sea slugs); they prey
on hydroids and coral polyps, and some even prey on venomous jellyfish. Some
species are able to digest the cnidarians without disturbing their cnidocytes,
and these cells are moved to the back of the nudibranch and protect it against
the nudibranch's own predators. The bright colors are though to be warning coloration.
The protuberances on the back are the "branch's" - the external gill-like
respiratory organs.
Snails
have put a 'twist' on molluscan evolution. The shell and mantle rotate during
development, placing the mantle cavity at the back of the animal. Many snails
are grazers; they scrape algae off rocks with a file-like tongue called a radula.There
are many predatory snails, however. Oyster drills have a tough radula, and they
scrape a hole through oyster shells. They then extend their proboscis through
the hole and consume the oyster. Coneshells, in the genus Conus, have
a modified radula that takes the shape and function of a harpoon. They stab
fish and inject a potent toxin; some Conus snail toxins are also lethal
to humans. Almost all molluscs are marine or aquatic; only some snails have
colonized land.The most coloful molluscs are nudibranchs (sea slugs); they prey
on hydroids and coral polyps, and some even prey on venomous jellyfish. Some
species are able to digest the cnidarians without disturbing their cnidocytes,
and these cells are moved to the back of the nudibranch and protect it against
the nudibranch's own predators. The bright colors are though to be warning coloration.
The protuberances on the back are the "branch's" - the external gill-like
respiratory organs.
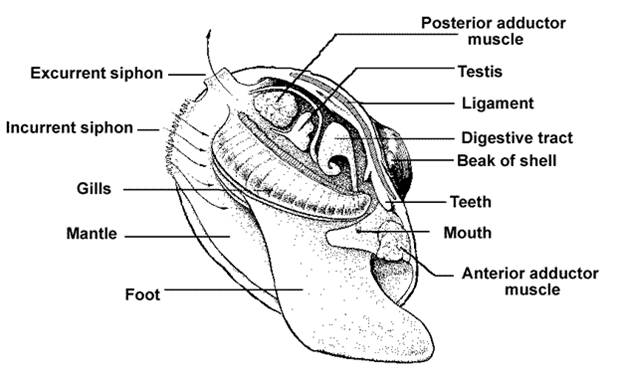 Bivalves
are an unusual group of molluscs. Bivalves are aquatic/marine filter feeders;
they lay on the sediment and filter water between the 'valves' of their shell.
They are bilaterally symmetrical; the midline of their back runs along the hinge
of their shell - so their shell comes down on both sides of the animal, covering
the animal and the foot. However, since the animals are sessile filter feeders,
their nervous system has evolved to be decephalized (decentralized). They don't
have a head - rather, because they do not move, their sensory systems have been
redistributed around the organism so they can perceive their environment from
all directions. Scallops, for instance, have eyes all around the margin of their
mantle and shell. So again, here is a sort of exception that proves the rule:
bilaterality directs a pattern of motion, which selects for cephalization at
one end. Some sessile organisms that don't move through the environment are
decephalized.
Bivalves
are an unusual group of molluscs. Bivalves are aquatic/marine filter feeders;
they lay on the sediment and filter water between the 'valves' of their shell.
They are bilaterally symmetrical; the midline of their back runs along the hinge
of their shell - so their shell comes down on both sides of the animal, covering
the animal and the foot. However, since the animals are sessile filter feeders,
their nervous system has evolved to be decephalized (decentralized). They don't
have a head - rather, because they do not move, their sensory systems have been
redistributed around the organism so they can perceive their environment from
all directions. Scallops, for instance, have eyes all around the margin of their
mantle and shell. So again, here is a sort of exception that proves the rule:
bilaterality directs a pattern of motion, which selects for cephalization at
one end. Some sessile organisms that don't move through the environment are
decephalized.
 The
last group of molluscs are the other-wordly cephalopods. Although they look
very different from a chiton, the evolution of their body plan entailed a rather
simple modification - the folding of the animal head to tail, and the elongation
of the back. The folding of the animal created an anterior half to the foot,
which evolved into tentacles, and a posterior half to the foot, which evolved
into the siphon. The mantle forms a muscular cowl over the organism, and water
can be sucked into the mantle cavity through the siphon, aerating the gills,
and then expelled by the muscular contraction of the mantle - propelling the
animal in a direction determined by the aiming of the siphon. As their name
implies, the cephalopods have a big head - and it is a brainy, smart head, too.
This correlates with their predatory behavior. As a general rule, predators
are pretty clever animals - at least more clever than their prey. This is especially
true for highly active and mobile predators like cephalopods, cats, and dogs.
Cephalopods are the largest living invertebrates, and they are also generally
regarded as the smartest. They can learn by mimicry. In a famous study, two
octopuses were placed in neighboring glass aquaria that had red and blue balls
at one end. One octopus had been trained to travel to the other end of the aquarium
and pick up a red ball to receive food. After observing this behavior, the second
untrained octopus would travel to the end of its aquarium and pick up a red
ball (not a blue one). That is alot of higher processing occuring. Within the
cephalopods, the evolutionary trend has been the reduction, internalization,
and loss of the shell. A shell is very heavy; reducing its size increases the
mobility of the predator. Internalizing the shell, as a rigid strut ("pen")
in squid allows it to help the hydrodynamics of the organism. For octopus,which
are sit and wait predators, the loss of the shell was adaptive because it meant
that octopus could squeeze into very narrow cavities to escape their predators
and to surprize their prey.
The
last group of molluscs are the other-wordly cephalopods. Although they look
very different from a chiton, the evolution of their body plan entailed a rather
simple modification - the folding of the animal head to tail, and the elongation
of the back. The folding of the animal created an anterior half to the foot,
which evolved into tentacles, and a posterior half to the foot, which evolved
into the siphon. The mantle forms a muscular cowl over the organism, and water
can be sucked into the mantle cavity through the siphon, aerating the gills,
and then expelled by the muscular contraction of the mantle - propelling the
animal in a direction determined by the aiming of the siphon. As their name
implies, the cephalopods have a big head - and it is a brainy, smart head, too.
This correlates with their predatory behavior. As a general rule, predators
are pretty clever animals - at least more clever than their prey. This is especially
true for highly active and mobile predators like cephalopods, cats, and dogs.
Cephalopods are the largest living invertebrates, and they are also generally
regarded as the smartest. They can learn by mimicry. In a famous study, two
octopuses were placed in neighboring glass aquaria that had red and blue balls
at one end. One octopus had been trained to travel to the other end of the aquarium
and pick up a red ball to receive food. After observing this behavior, the second
untrained octopus would travel to the end of its aquarium and pick up a red
ball (not a blue one). That is alot of higher processing occuring. Within the
cephalopods, the evolutionary trend has been the reduction, internalization,
and loss of the shell. A shell is very heavy; reducing its size increases the
mobility of the predator. Internalizing the shell, as a rigid strut ("pen")
in squid allows it to help the hydrodynamics of the organism. For octopus,which
are sit and wait predators, the loss of the shell was adaptive because it meant
that octopus could squeeze into very narrow cavities to escape their predators
and to surprize their prey.
Study Questions:
1. What is the benefit of a highly convoluted gut in flatworms?
2. Why are flatworms pre-adapted to life as a gut parasite? (Think about SA/V ratios).
3. List three types of annelids.
4. What is the advantage of a digestive tract, compared to a gut sac?
5. Why was segmentation - the replication of body units - such an important evolutionary innovation? (Think of the advantages of gene duplication.)
5. Draw the basic body plan of a mollusc, including the gut, foot, mantle, and gill. How is this modified to form a clam and a squid?
6. Why have clams "lost their head"?