 I.
Mendel's Contributions
I.
Mendel's Contributions
I.
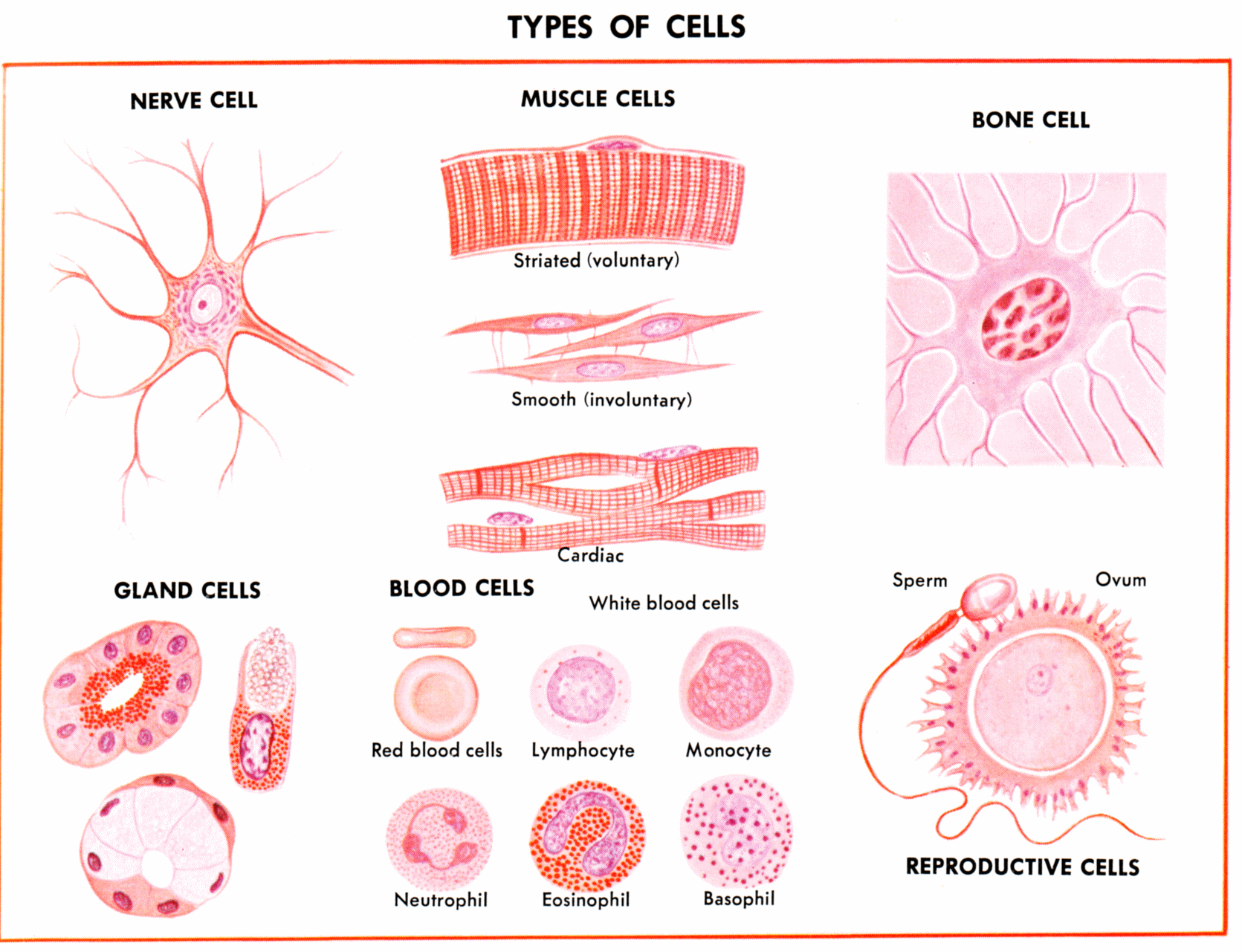
Mendel's ContributionsOne of the most remarkable things about multicellular life is the co-ordinated activity of cells. Think about it - you are sitting here "reading" - which is to say that light is impinging on cone and rod cells in your retina, electric impulses are created in these cells and passed to intermediary cells and to your optic nerve, and finally to the nerve cell bodies in the visual cortex in your brain. Here, through processes yet to be described, the emergent property of conscious thought makes sense of the pattern of excitation among these neurons and you interpret the perception of these black squiggles to mean something conceptually; a concept that you have encoded as a memory - probably as a particular combination of neuron cell firings in another part of your brain. At the same time, there are other electrical feedbacks between cells that are unconsciously adjusting muscle cells that regulate the aperture of your iris and the shape of your lens to maximize focus and visual acuity as you read. Red blood cells in your circulatory system are nourishing your brain with oxygen, propelled by the contraction of heart muscle cells. Glucose is in your blood, too; produced by the action of digestive enzymes secreted by other cells in your digestive tract, stored for a time in your liver, and released to your bloodstream under chemical cues from the cells in your pancreas. And other cells in you arm contract, causing you to unconsciously scratch the hair follicle cells on your head. Each of us - and every other complex multicellular organism - is composed of millions to trillions of cells that act in a co-ordinated way that maintains the lives of all of these cells and the organism they comprise. And whale cells are swimming through the oceans right now, doing whale things... and dragonfly cells are crawling through the mud at the bottom of the pond, doing dragonfly things. It's no wonder that it took about 3 billion years for life to evolve multicellularity - achieving a level of sophisticated communication and co-operation among microscopic and 'unconscious' cells is quite a feat! If this doesn't make you stop to consider how wonderful life is, then maybe biology isn't the right major for you!
Oh, and let's make it even more amazing. All of these cells that do such remarkably different things in your body are all THE SAME genetically. (This is a pretty important point: the differences in cell function DO NOT arise because different cells have different genes. Rather, all cells have ALL your genes, and different cells just read different subsets of genes and make differnt proteins.) You began your existence as a fertilized zygote - the fusion of your father's sperm and your mother's egg. This single cell divided by mitosis to form 2, then 4, then 8, then 16 cells. All these cells were identical, inheriting the complete genome from the zygote through these mitotic divisions. The process continued, with cells dividing by mitosis to produce the trillions of cells that comprise your body. Although somatic mutations occurred along the way, making some cells a little different, by and large all the cells in your body have the SAME genes. Again, these cells differ in shape and function because they read different parts of your genetic recipe book. Brain cells read recipes for neurotransmitters, and stomach cells read recipes for digestive enzymes, but BOTH neurons and stomach cells have genes for neurotransmitters and digestive enzymes. We belabor the point, but it is an essential fact that you must know from now on.
In this lecture, we are going to look at how genes get turned 'on' and 'off' in cells. We will use examples from bacteria and yeast, which are both single celled. However, by the logic presented above, you should see WHY these regulatory pathways, which function in much the same way in multicellular organisms, is so important to the history of life on earth and to your own personal existence.
Regulation reprise: We have already talked about gene regulation. At the end of the lecture on protein synthesis, there was a brief overview of how this process can be regulated. The accessibility of the gene can be regulated by histones, the binding and action of the RNA polymerase can be regulated by transcription factors, the m-RNA can be spliced differently in different cells to produce different products, the initial protein product can be spliced and/or modified differently in different cells to make different products, and environmental factors can influence how these things are done. Here, we are going to take a look at a couple of those processes in a bit more detail.
Some genes are on all the time in every cell - these are called constitutive genes. Can you think of genes that would have to be on in every cell? Well, all organisms need to be making phospholipids all the time, so I bet the genes that code for enzymes invovled in the production of phospholipids are on all the time. Genes for helicase and RNA polymerase, and for ribosomal proteins are probably on all the time in all cells, otherwise protein synthesis for the whole cell would shut down.
Other genes, however, are regulated. In some cases, the presence of another molecule turns the gene 'on' - these are inducible systems and the molecule that turns the gene on is called the inducer. Other molecules may turn a gene 'off' - this is a repressible system and the molecule that supresses gene activity is called a repressor. Inducers and repressors often bind to the DNA directly to exert their effects, usually upstream from the gene.
In E. coli bacteria, some genes that produce proteins involved in the same reaction are turned 'on' as a group - as a functional - or operational - unit. This unit is called an 'operon'. The lac operon is a functional unit of DNA that codes for three different enzymes involved in lactose metabolism - they influence the absoprtion of lactose and the splitting of lactose into glucose and galactose. So, this operon is used when glucose is needed by the cell and lactose is available. Curiously, these genes only get switch 'on' when lactose is present and glucose is absent - the exact environmental conditions under which the production of these genes that metabolize lactose and produce glucose would be adaptive. But how does this little bacterium know? How does it know that lactose is present and glucose is absent, and that it should turn on its genes? Well, it doesn't know. But bacteria that make these metabolic choices are strongly selected for, because the bacteria only make the enzymes when the enzymes are needed. This saves energy; energy that the bacteria can spend on other things like reproduction. (Selection is differential reproductive success...)
 1.
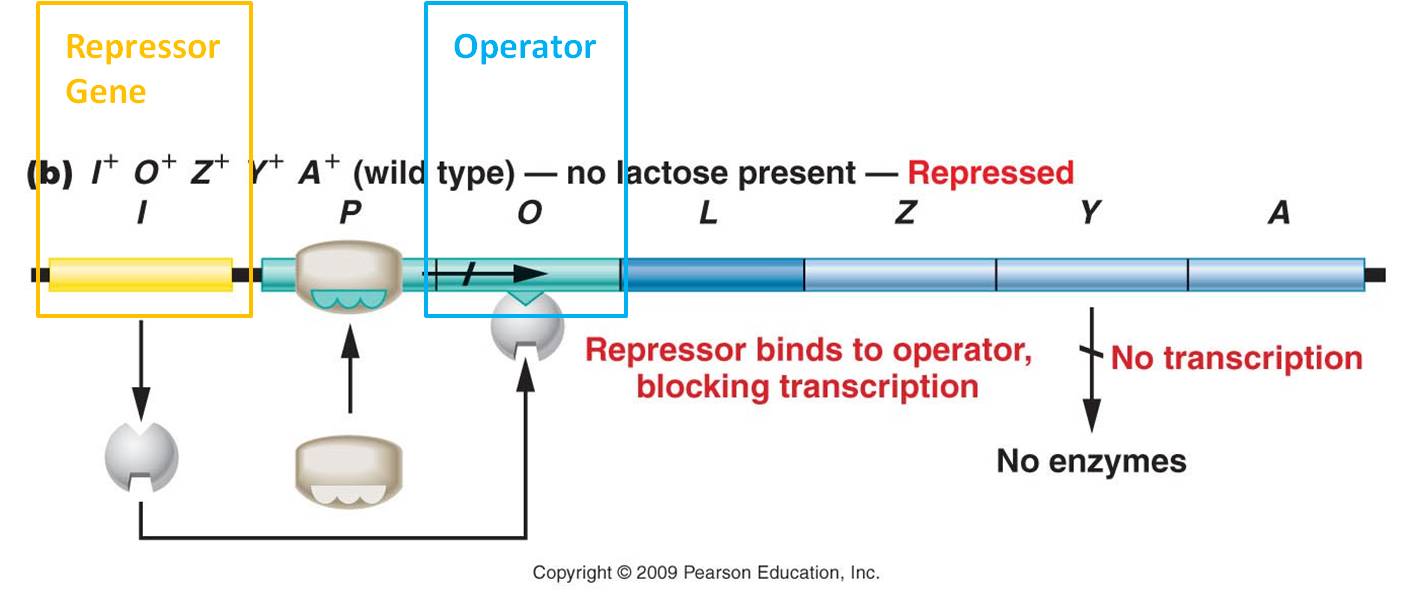
The structure of the operon
1.
The structure of the operon
- The three genes in the lac operon code for b-galactosidase (that cleaves lactose into glucose and galactose), permease (that increases the uptake of glucose), and transacetylase (also required for the process of lactose metabolism).
- Upstream from these genes is the Promoter, where the RNA Polymerase will bind before it transcribes the sense strand in these genes and makes the m-RNA product.
- Between the promoter and the genes is the 'operator' - the region where regulatory elements bind to disrupt or encourage transcription.
Elsewhere in the genome is the repressor gene, which codes for the repressor protein.
 2.
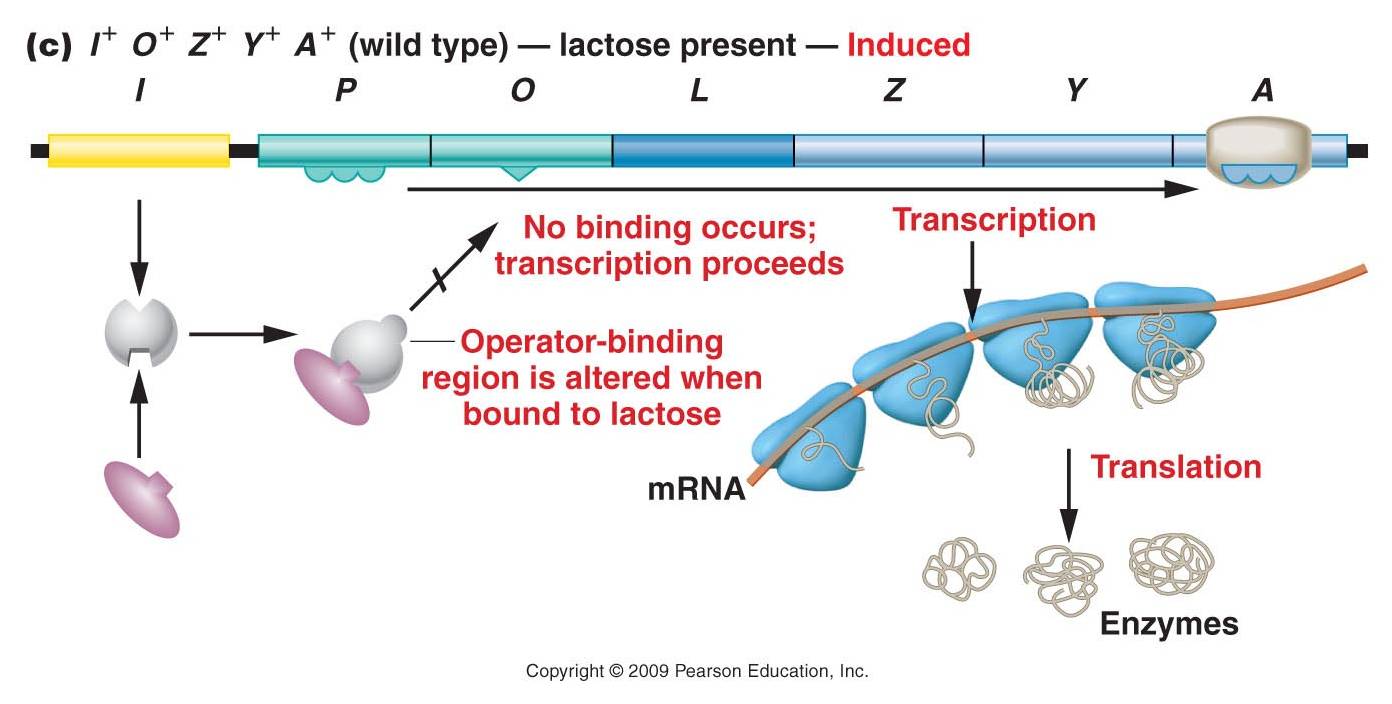
Induction of the lac operon by lactose
2.
Induction of the lac operon by lactose
Jacob and Monod constructed this operon model of gene regulation in 1960, and they hypothesized that it was an inducible system. The repressor gene produced the repressor protein that, in the absence of lactose, binds to the operator region in the operon. The binding of this protein blocks the RNA polymerase from transcribing the genes, so the genes are 'off'. In the presence of lactose (the inducer), lactose binds to the repressor and changes the repressor's shape. With the new shape, the repressor can no longer bind to the operator and block RNA polymerase. So, RNA polymerase transcribes the genes and the enzymes for lactose metabolism are produced. Eventually the lactose levels decline, the inhibition on the repressor is released, and the repressor can bind again to the operator and stop transcription. So, the enzymes that are used to metabolize lactose are only produced when lactose is present. This is a very adaptive, energetically efficient system.
 Mutant
analyses confirmed the model. A mutation in the operator changed the binding
site for the repressor, and the repressor couldn't bind. so, the genes were
'on' all the time, even in the absence of lactose. Changes to the repressor
gene caused two effects. Changes to the operator binding site meant that it
couldn't bind the operator and couldn't block transcription. Changes to the
lactose binding site meant that it would ALWAYS bind the operator because it
would not bind lactose and change shape.
Mutant
analyses confirmed the model. A mutation in the operator changed the binding
site for the repressor, and the repressor couldn't bind. so, the genes were
'on' all the time, even in the absence of lactose. Changes to the repressor
gene caused two effects. Changes to the operator binding site meant that it
couldn't bind the operator and couldn't block transcription. Changes to the
lactose binding site meant that it would ALWAYS bind the operator because it
would not bind lactose and change shape.
3. Repression of the lac operon by Glucose
There are two sites in the promoter; a site for the RNA polymerase and another site called the 'catabolic activating protein' site (or 'CAP' site). The CAP is a transcription factor - binding of this protein to the promoter greatly enhances the binding of the RNA polymerase to the promoter. However, CAP will only bind if c-AMP concentrations are high, and THIS only occurs if glucose concentrations are low! So, if [glucose] is high, c-AMP is low, and the CAP doesn't bind to the CAP site. RNA polymerase doesn't bind to the promoter, and transcription of these genes does not occur - regardless of the concentration of lactose. This is adaptive; glucose is what the cell needs. If glucose is present, then there is no benefit to metabolizing lactose to make glucose - so energy is saved by keeping these genes off. If [glucose] is low, then c-AMP is high, CAP is activated and binds to the CAP site, enhancing binding of the RNA polymerase. So, the operon is ready to go, and if lactose is present, then the repressor will not bind to the operator and the polymerase can transcribe the genes. In this way, the lactose will be metabolized and glucose will be produced.
 D.
Summary
D.
SummaryThe regulation of the lac operon
is interesting, because it shows how other genes in the genome (repressor gene)
and a combination of environmental variables (lack of glucose, presence of lactose)
affect the expression of a gene. It is both complicated - in that it is an adaptive,
dynamic response to changes in the environment, and simple - in that the mechanisms
involve a small number of proteins acting in a rather simple and direct way
(literally blocking RNA polymerase, in some cases). Many genes are affected
by environmental cues from beyond the cell: these can be nutrients (like glucose
and lactose), light energy (day length turns genes for flowering on in many
plant species), and hormones (estrogens and testosterones secreted by endocrine
glands during adolescence turns on genes that co-ordinate sexual development
throughout the body), to name a few. The more regulators there are, the more
refined and subtle the response can be - like any polygenic trait. In addition,
the more regulators there are, the more pathways there are to activate the gene.
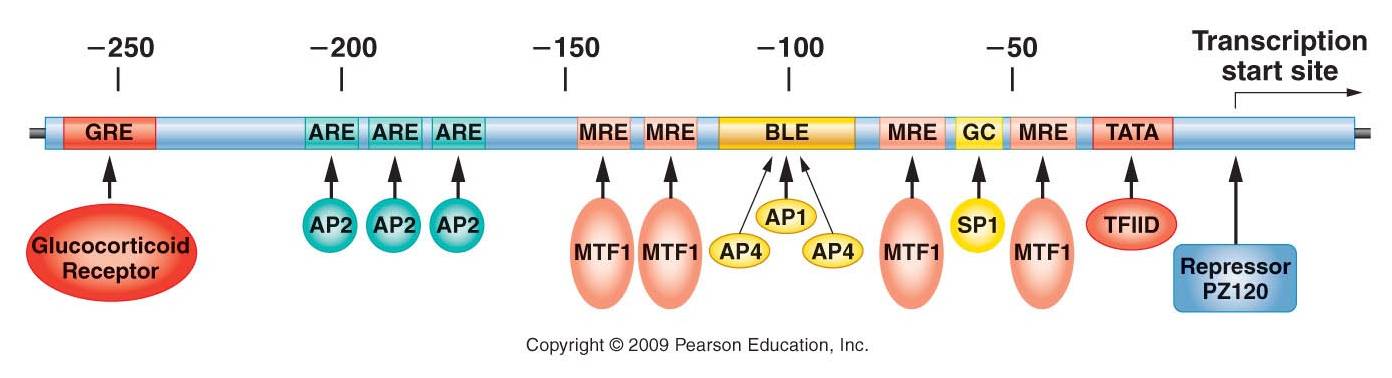
 Look at the picture,
at right. These are the binding sites of all the transcription factors that
can affect the expression of the human metallothionien IIA gene. So, the production
of this protein can be stimulated or inhibited by a variety of factors. The
protein is involved in detoxification of metals, but it is also involved in
axon elongation after neuron damage. These two functions are completely independent,
and so it has to be turned on by different cues related to each function.
Look at the picture,
at right. These are the binding sites of all the transcription factors that
can affect the expression of the human metallothionien IIA gene. So, the production
of this protein can be stimulated or inhibited by a variety of factors. The
protein is involved in detoxification of metals, but it is also involved in
axon elongation after neuron damage. These two functions are completely independent,
and so it has to be turned on by different cues related to each function.
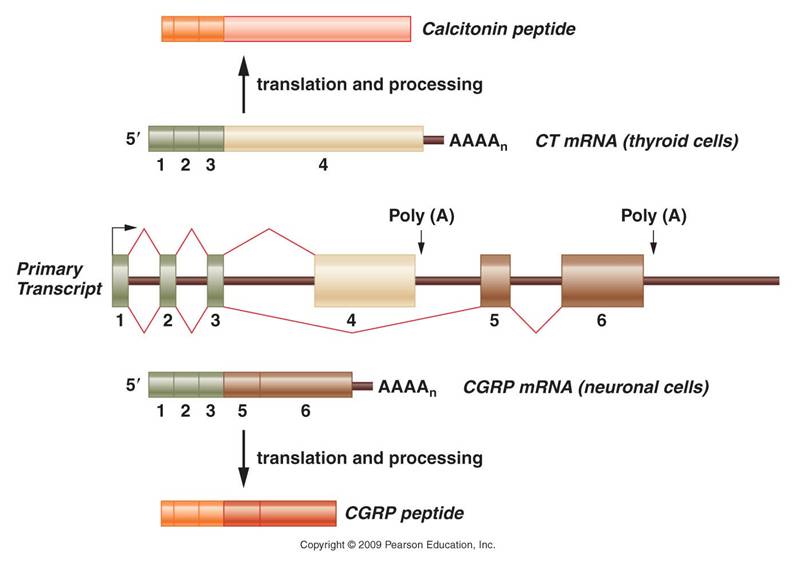
We also discussed the ability of single genes to encode different protein products. One way this happens is by splicing together different exons in different cells. Here is an example, in the picture to the right. The gene has six exons. When transcription occurs, an intial m-RNA product is produced that has 6 exons, too. In neurons in the brain, this m-RNA is spliced in such a way that exon #4 is not incorporated in the functional m-RNA; the functional m-RNA that gets translated contains exons 1,2,3,5, and 6. The protein that is made is a brain hormone. In the thyroid gland (in your neck), however, this same initial m-RNA transcript is spliced a different way, and includes only exons 1,2,3 and 4. This protein is a calcium regulator in the thyroid. So, one gene codes for two different proteins in different cells - not because of DNA differences (the gene is THE SAME), but because of how the initial m-RNA product is spliced.
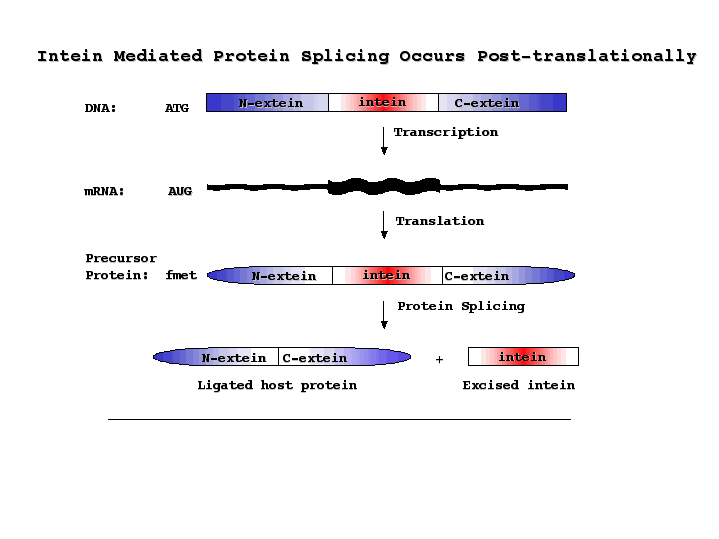
Finally, there is post-translational processing. This can be protein splicing, where regions are cut out in differnt ways in different tissues. In this case, the terminal portions, called "extiens" (like exons, but in a protein) are joined together after the excision of the "intein" (like intron).
Gene regulation is an extraordinarily complex and simple process. Each regulatory pathway is simple, but multiple pathways create a complexity and subtlety that allows for the specialization fo structure and function that makes multicellular life possible. That makes it cool.
 Study
questions:
Study
questions:
1. Describe the effects of glucose and lactose concentrations on the operation of the lac operon. How is this adaptive?
2. The human genome contains about 30,000 genes, but produces over 100,000 proteins. How is that possible?