 A.
Overview
A.
Overview
Meiosis produces specific types of reproductive cells. In animals, meiosis occurs in the gonads (ovaries/testes) and produces eggs or sperm. In plants and fungi, meiosis produces other reproductive cells called spores. The production of egg and sperm occurs for only one purpose - sexual reproduction. Sperm and egg must fuse (fertilization) before the development of the offspring can begin. Spores do not fuse; a spore is a single reproductive cell that simply begins to divide and grow on its own to become an offspring. In this lecture, we will ask the question "why sex?" Of course, in an evolutionary context, this question means "is this complex trait adaptive, and if so, how?" After describing the adaptive significance of asexual and sexual reproduction, we will describe the process, its relevance to Mendel's principles of heredity, and we will solve Darwin's dilemma regarding the source of heritable variation.
A.
OverviewWe have already considered the broader of issue of reproduction, itself. And we have examined one type of reproduction - mitosis. Although mitosis is the way that single eukaryotic cells divide, it is important to appreciate that, for single-celled eukaryotes (protists), this is a mechanism of organismal reproduction, as well. In addition, many organisms that are multicellular use mitosis as a form of organismal reproduction. For example, many plants grow lateral 'runners' along the ground or under the soil. From the end of this runner, a new bud can develop that grows roots and shoots. This bud can grow into an individual plant that is capable of living on its own, even after the runner is cut. All the cells in this new independent plant were produced by mitotic divisions from the parent plant. Indeed, this new plant is genetically identical to the original plant - it is a 'clone'. Fragmentation is another example of clonal reproduction in plants, where a branch that is broken or cut from a plant takes root and becomes in independent organism. This is more common for aquatic plants, where the broken fragment can stay alive and not dry out while it grows its own roots. Clonal reproduction also occurs in many animals. Corals and Hydroids (like the Hydra pictured at right) will grow a new polyp by mitotic division, and this polyp can 'bud off' the parent and become a new independent organism. In these cases, an offspring has been produced by a parent through mitosis, alone. The offspring is genetically identical to the parent. This clonal form of reproduction is also called "asexual reproduction".
Sexual reproduction represents the formation of a new genotype. This occurs in a number of ways in nature. In bacteria, genes can be donated from one cell to another, or even absorbed from the environment. This changes the genotype of the bacterium the receives these new genes. A new genotype has formed, even though there is no no organism produced. Many protists have multiple nuclei, and they exhibit a form of sex by exchanging micronuclei during conjugation. This also forms new genotypes without forming new organisms.
 Typically,
however, sexual reproduction occurs through the fusion of specialized reproductive
cells called gametes. In some organisms (like fungi and some algae), the gametes
are all about the same size and are usually motile (with flagella). These species
are called isogamous (iso = equal). These species
don't really have 'males' or 'females'. Rather, individuals belong to 'mating
types' that produce genetically different types of gametes. Indeed, in most
of these species, there are many 'mating types' or 'sexes', defined by the capacity
of their gametes to fuse with other mating types (while not being able to fuse
with their own mating type). Isogamy is thought to be the ancestral form of
sexual reproduction. There are several theories that explain how a two sex (male/female)
mating system could have evolved from this multiple mating-type ancestor. The
first step was probably selection for one gamete to become larger while the
other became smaller, resulting in anisogamous species
with two distinctly different sized gametes that may still both have flagella.
From this stage, oogamy evolved, in which the larger
gamete became an unflagellated egg. In hermaphroditic
species (like many plants and invertebrate animals, and a few rare deep-sea
fish), single individuals produce both sperm and egg in different organs. In
plants, hermaphrodism describes plants with 'perfect' flowers, containing both
male and female organs. In some plant species, individuals have separate male
flowers and female flowers - these are monoecious
species. If a single organism can produce both sperm and egg, then it might
have the capacity to 'mate with itself'. These organisms are called 'self-fertile'.
Although mating with one's self makes searching for a mate rather easy, there
are some genetic downsides that we will consider in more detail later. So, many
of these species have evolved genetic mechanisms that prohibit an individual's
sperm from fertilizing its own eggs. These organisms are called 'self-incompatible'.
Even without a genetic incompatibility, many organisms reduce the changes of
self-fertilization by releasing their sperm and egg at different times.
Typically,
however, sexual reproduction occurs through the fusion of specialized reproductive
cells called gametes. In some organisms (like fungi and some algae), the gametes
are all about the same size and are usually motile (with flagella). These species
are called isogamous (iso = equal). These species
don't really have 'males' or 'females'. Rather, individuals belong to 'mating
types' that produce genetically different types of gametes. Indeed, in most
of these species, there are many 'mating types' or 'sexes', defined by the capacity
of their gametes to fuse with other mating types (while not being able to fuse
with their own mating type). Isogamy is thought to be the ancestral form of
sexual reproduction. There are several theories that explain how a two sex (male/female)
mating system could have evolved from this multiple mating-type ancestor. The
first step was probably selection for one gamete to become larger while the
other became smaller, resulting in anisogamous species
with two distinctly different sized gametes that may still both have flagella.
From this stage, oogamy evolved, in which the larger
gamete became an unflagellated egg. In hermaphroditic
species (like many plants and invertebrate animals, and a few rare deep-sea
fish), single individuals produce both sperm and egg in different organs. In
plants, hermaphrodism describes plants with 'perfect' flowers, containing both
male and female organs. In some plant species, individuals have separate male
flowers and female flowers - these are monoecious
species. If a single organism can produce both sperm and egg, then it might
have the capacity to 'mate with itself'. These organisms are called 'self-fertile'.
Although mating with one's self makes searching for a mate rather easy, there
are some genetic downsides that we will consider in more detail later. So, many
of these species have evolved genetic mechanisms that prohibit an individual's
sperm from fertilizing its own eggs. These organisms are called 'self-incompatible'.
Even without a genetic incompatibility, many organisms reduce the changes of
self-fertilization by releasing their sperm and egg at different times.
 In
dioecious (two 'houses') species, individuals are
either male or female. Males are defined as the sex that produces sperm, while
females are defined as the sex that produces the large egg. This is a more comprehensive
way to define a sex, allowing us to define sexes in species where the sexes
are morphologically indistinguishable. Many fish, for instance, lack genitalia
and simply excrete their gametes - egg or sperm - into the water for fertilization.
In some species, individuals change their sex as they develop and age. This
is called sequential dioecy or sequential
hermaphrodism. Organisms that are males first and then change
to females are called protandrous, while individuals
that are females first and then change to males are called progynous.
In
dioecious (two 'houses') species, individuals are
either male or female. Males are defined as the sex that produces sperm, while
females are defined as the sex that produces the large egg. This is a more comprehensive
way to define a sex, allowing us to define sexes in species where the sexes
are morphologically indistinguishable. Many fish, for instance, lack genitalia
and simply excrete their gametes - egg or sperm - into the water for fertilization.
In some species, individuals change their sex as they develop and age. This
is called sequential dioecy or sequential
hermaphrodism. Organisms that are males first and then change
to females are called protandrous, while individuals
that are females first and then change to males are called progynous.
As you can see from this introduction, sex can be alot more complicated than you might have thought. It can involve the production of new organisms, or only new genotypes. It can involve multiple sexes, or just two sexes. It can involve organisms that are one sex, two sexes, or change sex, and organisms that mate with themselves. However, even given all this variation, the common element in every case of sexual reproduction is the production of a new genotype. In asexual reproduction, the daughter cells are nearly identical to the parent cell (except for rare mutations). Next, we will compare the adaptive benefits of each type of reproduction. And remember, adaptations are traits that increase the reproductive success of a genotype in a given environment.
 1.
The Costs and Benefits of Asexual Reproduction:
1.
The Costs and Benefits of Asexual Reproduction:
a. Benefits:
1) Probability of mating is high - you don't need to find a mate!
2) All else being equal, reproductive output and the transmission of genetic information to the next generation should be high. In fact, asexual reproduction maximizes the transfer of parental genes to the next generation: every offspring receives the entire genome of the parent. So, in terms of "differential reproductive success" (what's that? - you'd better know!!!), asexual reproduction has a quantitative edge in getting parental genes into the next generation.
3) In a constant environment, the offspring should all do well (they have the parent's genome, and the parent survived and reproduced in this environment with that genome). So, when the environment is stable, the probability of offspring survival and reproduction should be high. So, in a constant environment, clonal/asexual organisms should reproduce easily and quickly, and their offspring should thrive and reproduce.
b. Costs:
1) Mutations occur, and most are deleterious (have negative effect). From one generation to the next, these mutations will accumulate in a lineage because every parental gene is passed to the offspring. There is no way to "get rid" of bad genes or deleterious mutations.... this is called "Muller's Ratchet". Selection can weed out bad genes from a population, but over time all lineages will be burdened by the accumulation of deleterious genes.
2) Mutations become the only source of beneficial, adaptive variation. Because mutation rates are low, reproduction must be very rapid (prodigious, like bacteria), to create enough variation through mutation, alone, to by chance produce something new and useful.
3) Few environments on Earth are stable over the long term. When an environment changes, survival of offspring will be an "all or none" affair. If that genome can't survive in the new environment, then all the offspring die and that lineage comes to an end. Over time, we might expect every lineage in a species to encounter an environment to which it is not suited, resulting in the extinction of the species.
2. The Costs and Benefits of Sexual Reproduction:
a. Costs:
1) For organisms that cannot mate with themselves, sexual reproduction can be an energetically expensive behavior. There are energetic costs to finding and acquiring a mate.
2) Only 1/2 a parent's genes are passed to each offspring; so it is not as effective at transmitting parental genes to the next generation as asexual reproduction.
3) Many offspring will inherit combinations of genes poorly suited for the environment - or at least more poorly suited than the parental combination. In the luck of the draw, some offspring may receive most of the parents' 'bad' genes.
b. Benefits:
1) Since each offspring inherits only 1/2 the genes from each parent, deleterious genes can be purged from a lineage just be chance. In other words, just by chance, many of the offspring will have 'lucked-out' and will not inherit any of the new, deleterious mutations produced in the preceeding generations. This eliminates the effects of Muller's ratchet.
2) By combining genes from two parents, or by exchanging genes between organisms, new variation is produced MUCH more rapidly than by mutation, alone. Although many of these new genetic combinations may perform worse than the parents in a given environment, some may perform better. Better combinations will be able to harvest energy and convert it into offspring more effectively. In other words, "better" means better able to survive and reproduce in an environment.
3) When the environment changes (and it almost always will), it is more likely that, among the variable genomes produced by sexual reproduction, there will be a genome that can tolerate (if not excel) in this new environment. So, over the long term in changing environments, asexual lineages are likely to meet an environment they can't tolerate and go extinct, while sexual lineages are more likely to persist.
3. Summary:
Many organisms reproduce asexually. Indeed, for many organisms like bacteria, asexual reproduction is the primary mode of reproduction. Asexual reproduction is adaptive when environmental conditions are stable; it is a highly effective way for a genotype or lineage to 'out reproduce' other genotypes or lineages in the environment, and acquire limiting resources. However, because sexually reproducing species produce more variation per unit time, sexually reproducing species will adapt more rapidly to environmental conditions. In a changing environment, this means that sexually reproducing species can continue to adapt with the changes in the environment, while asexual lineages go extinct as they meet up with environments that they can't tolerate. The exception proves the rule here. If we look at organisms that reproduce asexually most of the time, we can ask "under what conditions do they switch to a sexual mode of reproduction?" The answer is usually: "when the environment changes." It is precisely under changing conditions that sexual reproduction is adaptive.
 1.
How? - Reduction and Fertilization
1.
How? - Reduction and Fertilization
There is a big benefit to combining genes from different organisms - new variations are produced that may be adaptive. Some organisms donate genes between cells, others trade nuclei. But for most sexually reproducing organisms, this mixing of genomes occurs through the production of specialized cells called gametes that fuse to form a new genotype and a new organism.
There is a problem to fusing normal body cells to produce this variation. By fusing body cells, the number of chromosomes and the amount of genetic information doubles each generation. As we'll see, many of the chemical reactions and patterns of genetic regulation depend on a constant balance of enzymes and substrates, and a constant balance of particular proteins. Doubling the genetic information can disturb these quantitative relationships. In addition, fusing body cells does not escape the constraints of Muller's ratchet - all genes are passed to offspring and the offspring now acquire all the bad genes from BOTH parents.
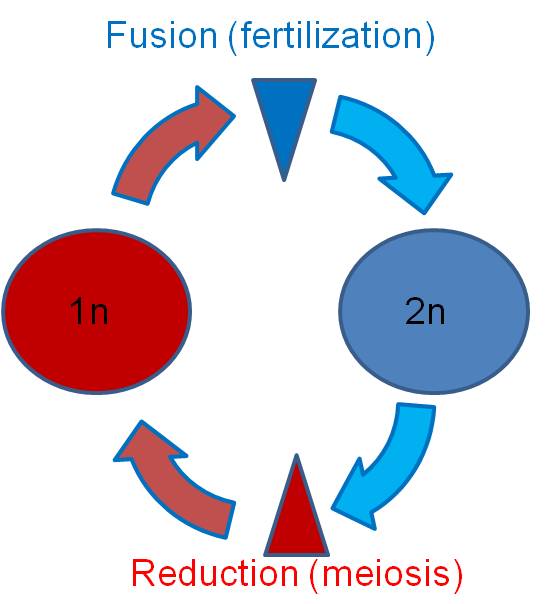
The solution that life evolved is a new process of cell division - meiosis. In this process that produces specialized reproductive cells, the genetic information is halved. This halving is also called reduction. If the cells produced by this process are gametes (remembering that plants and fungi use meiosis to produce spores), then the fertilization of sperm and egg reconstitutes the appropriate set of genetic information for that species.
2. When? - Life Cycles of Sexually Reproducing Organisms
The production of cells by meiosis and the fusion of cells to create new combinations occurs at different times in the life cycles of different organisms.
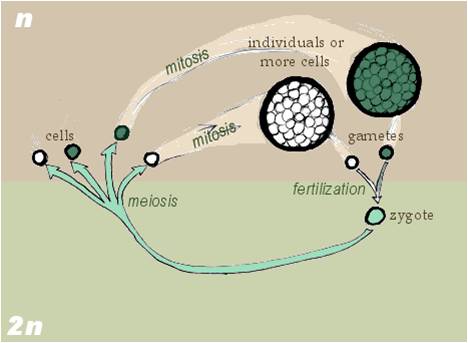
Zygotic meiosis: Most fungi (and many protists and green algae) are haploid. So, when they produce gametes, they produce them by mitosis. These gametes (or specialized nuclei) fuse during fertilization, producing a diploid cell. This zygote then divides by meiosis - producing haploid cells (spores) that - in multicellular representatives like the fungi - divide by mitosis and produce the multicellular haploid organism. |
 |
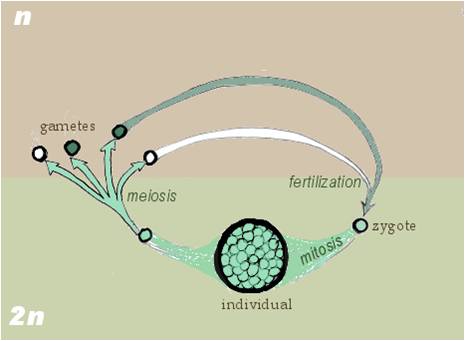
Gametic meiosis: Other organisms, like all animals, some fungi, and some brown algae, have diploid organisms. In these organisms, specialized cells in the multicellular organism divide by meiosis, producing haploid gametes. These gametes must fuse in a fertilization event. The resulting zygote divides mitotically, to produce the multicellular diploid organism. |
 |
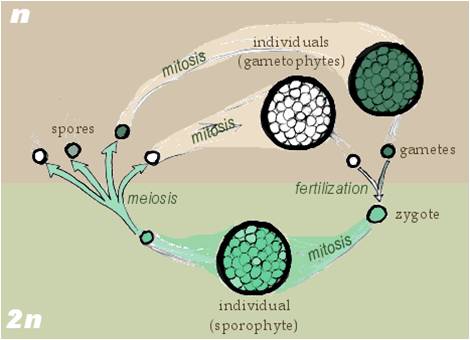
Sporic meiosis: Most plants and some fungi have yet another life cycle called alternation of generation - meaning that there are alternating generations of multicellular haploid organisms and multicellular diploid organisms. If we begin with the multicellular diploid organism, there are specialized cells that divide by meiosis, producing haploid reproductive cells. However, these are spores - not gametes - and these cells divide mitotically to produce the multicellular haploid offspring. In the multicellular haploid offspring, gametes (egg and sperm) are produced by mitosis. The sperm fertilizes the egg, producing a diploid zygote. The diploid zygote divides by mitosis, producing the multicellular diploid offspring of these multicellular haploid parents. So, the ploidy level alternates each generation. |
 |
1. Overview
of the Process:
Meiosis has two cycles of division, preceeded by an interphase containing a
G1, S, and G2 phase. So, the cell entering meiosis has replicated chromosomes.
The first cycle is called Meiosis I, or the reduction cycle. A diploid
cell containing two sets of replicated chromosomes divides and produces cells
that are haploid, containing one set of replicated chromosomes. The second cycle
is Meiosis II, also called the division cycle. In this cycle, the haploid
cells divide in a manner much like mitosis. In other words, the sister chromatids
of each replicated chromosome are separated, but there is no change in ploidy.
The haploid cell with replicated chromosomes divides into haploid cells with
unreplicated chromosomes .
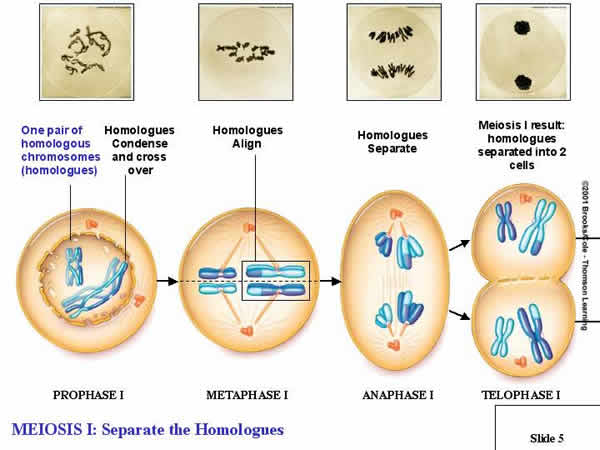 2.
Meiosis I (Reduction)
2.
Meiosis I (Reduction)
a. Prophase I:
- The chromosomes condense as paired homologs.
DEFINITION: Homologous chromosomes govern the same suite of traits, but may affect those traits in different ways. So, if there is a chromosome with a gene for purple flower color at a particular spot (locus), then it's HOMOLOG will also have a gene for flower color at that same spot... but it might be a different ALLELE for flower color (white).
- As condensation occurs, replicated, homologous chromosomes pair up. Each replicated chromosome has two chromatids, so the pair of homolos for a structure with four chromatics - a 'tetrad'. Piece of chromosome can be exchanged between homologs. This is called 'crossing over' . This creates new combinations of genes on chromosomes. we will consider this in more detail in a couple days.
- Asd the spindles join to paired homologs, the spindle fibers from each pole can only attach to ONE homolog or the other.
b. Metaphase I:
- THIS IS THE MOST IMPORTANT STEP, AND THE THING THAT DISTINGUISHES MEIOSIS FROM MITOSIS... Homologous pairs line up on the metaphase plate (NOT in single file as in mitosis, but as PAIRS).
c. Anaphase I:
- Whole, replicated chromosomes are drawn to each pole by the spindle fibers. So, if the 'A' chromosome goes to one pole, the 'a' chromosome goes to the other.
d. Telophase I:
- The cytoplasm divides; each new cell has only ONE chromosome from each homologous pair. If the cell started with 4 chromosomes (2n = 4), NOW each daughter cell has only 2 chromosomes (1n = 2). This is REDUCTION, and it is the most important part of Meiosis. It occurs because homologs pair up in metaphase I, rather than lining up in single file as in mitosis.
3. An optional interphase:
The transition from meiosis I to meiosis II cay vary between species. In some species, the nuclear envelope reforms around the chromosomes in each nucleus. There may even be an interphase in which the chromosomes decondense, protein synthesis occurs, and the cells grow. However, no S phase occurs in this interphase - the chromosomes are already in their replicated state. If this interphase does occur, then Prophase II begins with the condensation of the chromosomes and the breakdown of the nuclear membrane.
In other species, the cells proceed directly from Telophase I to Prophase II; the chromosomes are already condensed and the nuclear membrane is already broken down. In this case, the only thing that happens is that a new spindle forms, attaching each chromosome to both poles of the cell.
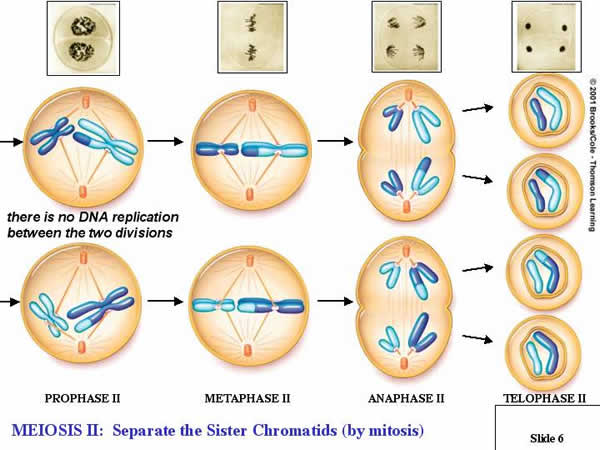 4.
Meiosis II: (Division):
4.
Meiosis II: (Division):
a. Prophase II:
Depending on the transition, events proceed such that the new spindle apparatus attaches condensed chromosomes to both poles of each cell produced in the reduction cycle (Meiosis I).
b. Metaphase II:
Replicated chromosomes line up in single file on the metaphase plate, in the middle of each cell.
c. Anaphase II:
The chromatids in each replicated chromosome are drawn to opposite poles in each cell.
d. Telophase II:
The nuclear membranes reform while cytokinesis divides the cells.
5. Modifications in Anisogamous Species:
In isogamous species and in homosporous plants (that produce one type of spore by meiosis), the reproductive cells produced by meiosis are all the same size. In this case, the process works as described above, and one parental cell produces four equal-sized daughter cells.
In anisogamous species (and in heterosporous plants that produce different sized spores by meiosis), the reproductive cells produced by male and female organisms vary in size. Although the division of the genetic material occurs in the same way (creating daughter cells that a have exactly half of the genetic information of the parent cell), the division of the cytoplasm differs in these processes.
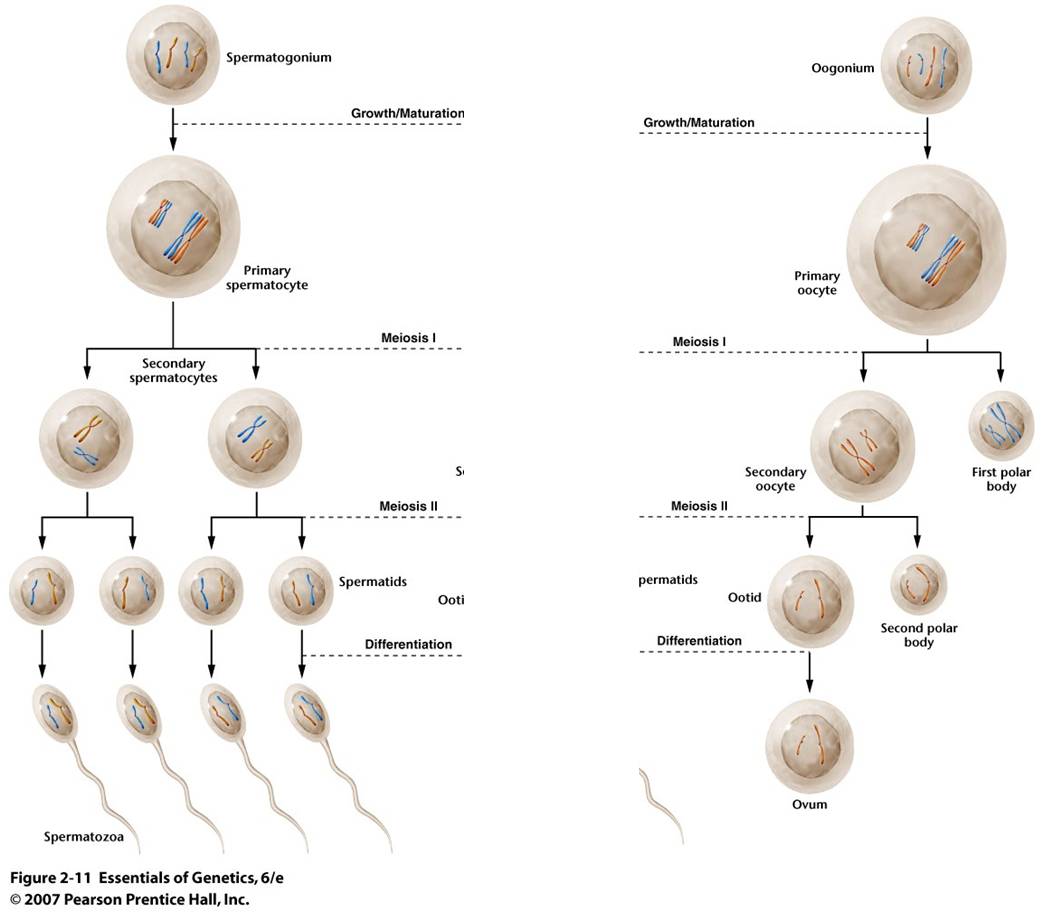 a.
Spermatogenesis:
a.
Spermatogenesis:
TIn sperm production, the division of the genetic information is EVEN, and the division of the cytoplasmic material is EVEN. So, Spermatogenesis produces four small functional sperm cells from one initial diploid cell.
b. Oogenesis:
Egg production is a little different. The division of the genetic information is EVEN, just as above. However, one of the cells produced in each divisional cycle receives almost ALL of the cytoplasm. So, after Meiosis I, there are two haploid cells, one is very small and the other is very large, having received almost all the cytoplasm. In fact, the small cell may be so small that it doesn't even have enough energy to divide again. The big cell DOES complete Meiosis II, but the cytoplasm is unequally divided again. So, another small cell is produced, and there is only one large functional egg cell produced. The small, non-functional cells are called "polar bodies." In many species, like humans, oogenesis stalls in prophase I. Girls are born with their full complement of egg-producing cells arrested in prophase I. With the onset of puberty, one cell each month completes the meiotic cycle and ovulation (eruption of the single egg from the ovary) occurs. This process of ovulating one egg per month continues throughout a woman's life until menopause. As such, many cells wait several decades before they complete meiosis. It is thought that this delay may increase the likelihood of divisional errors.... the longer a cell waits around, the more likely it is that when it divides it will not divide correctly. This would explain the increased frequency of genetic anomalies with increasing maternal age.
 1.
Meiosis and Mendelian Heredity
1.
Meiosis and Mendelian Heredity
Meiosis was first described in 1876 by Oscar Hertwig; the movement of chromosomes in meiosis was first described by Eduard Van Beneden in 1888. Sutton and Boveri, working independently in 1902, saw pairs of chromosomes separating in Meiosis I. Being familiar with the recently rediscovered work of Gregor Mendel, they appreciated this correlation between the movement of chromosomes during gamete formation and Mendel's principle of segregation. If one homolog carried one allele for a trait while the other homolog carried the other allele, then the separation of homologs could explain the segregation of alleles during gamete formation. Likewise, if the way that one pair of homologs separated had no effect on the way that other pairs of homologs separated, then the movement of chromosomes could explain the principle of independent assortment, as well.
They hypothesized that these chromosomes, moving in a pattern consistent and correlated with Mendel's two principles, might contain the heredity determinants. This is a big deal - it is called the "Chromosomal theory of heredity". Why do we know that chromosomes carry the genetic info? Because in 1902, Sutton and Boveri observed the correlation between the movement of chromosomes and Mendel's laws of heredity, and they proposed the hypothesis that chromosomes might carry the heredity information and cause patterns of heredity. This hypothesis was confirmed by Thomas Hunt Morgan's group working with fruit flies in 1915. Experiments in 1944 determined that it was the nucleic acid in chromosomes, and not the proteins, that was the hereditary information.
2. Solving Darwin's Dilemma:
Darwin observed that every population expressed considerable variation in every generation. He was confused by this, because if selection had been operating for millennia, why hadn't selection honed a species to the point where only the best adapted phenotype persisted and reproduced? In addition, Darwin was a supporter of the idea of blending heredity. This would further reduce variation each generation (white and black to grey...). However, because the hereditary material is PARTICULATE and not a blending fluid, sexual reproduction produces unique combinations of particles in EVERY generation. Indeed, the amount of new combinations that sexual reproduction can produce - in other words, the amount of genetic variation that sexual reproduction can produce - is staggering.
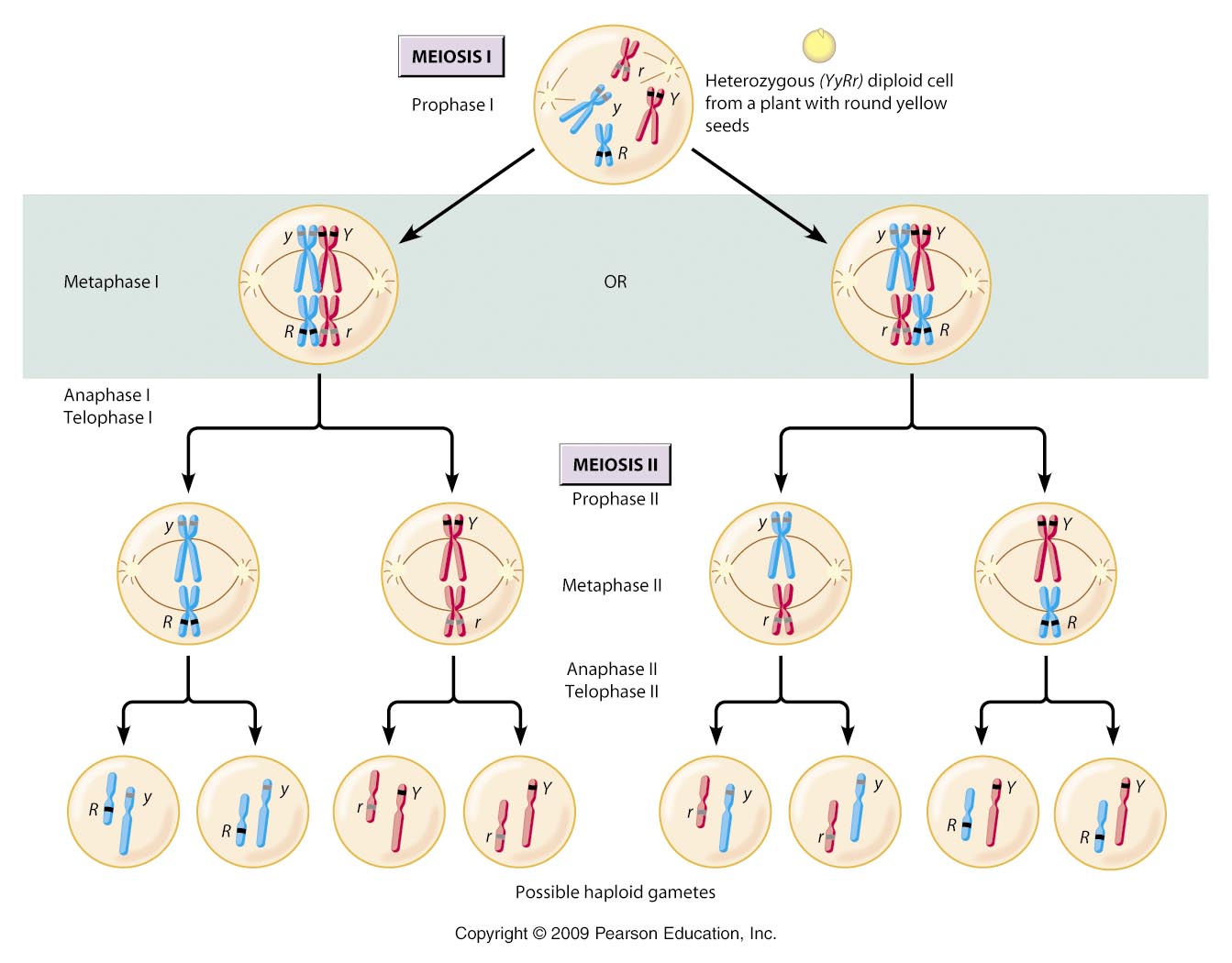
So, a cell 2n = 4, has two pairs of homologus chromosomes. Let's call one pair 'A' and 'a' and the other pair 'B' and 'b'.
***Don't get confused by thinking that these letters represent single genes; they do not. Rather, they simply represent different chromosomes in a homologous pair. Because chromosomes carry 100's - 1000's of genes, it is almost impossible for homologs to be genetically identical over their entire length. So, although two homologs might carry the same gene at a particular locus (which we could, confusingly, represent as 'AA'), the homologs will NOT be the same across all the genes they carry. So, we represent their similarity as homologs by using the same letter, but their necessary DIFFERENCE with capital and lower case versions of that letter. OK?***
Now, you know that this organism can produce 4 gametes as a consequence of segregation and independent assortment: AB, Ab, aB, ab.
A cell that is 2n = 6 has three pairs of homologs, AaBbCc. And, it can produce 8 types of gametes, ABC, abc, Abc, aBC, ABc, abC, AbC, aBc. So, there is a pattern here. The number of chromosomal combinations that are possible in the gametes = 2n (two to the 'n' power, where 'n' = the number of chromosomes in the haploid set). So, for 2n = 4, n = 2, and gamete number = 22 = 4. And, for 2n = 6, n = 3, and gamete number = 23 = 8.
Well, most organisms have lots of chromosomes... humans have 2n = 46. So, n = 23, and the number of different types of genetic combinations that we EACH can make in our gametes is 223 = about 8 million. That's more than the population of Georgia or New Jersey. That is a staggering amount of genetic variation, produced by each single human being. But of course, one gamete does not make an offspring; you need two to tango (so to speak). So, if a male can produce 8 million different types of sperm, and if a female can produce 8 million different types of eggs, and if any of the sperm are just as likely to fertilize any of the eggs, then there are 8 million x 8 million possible combinations of sperm and egg (that's one big Punnett Square!); that's 223 x 223 = 246: about 70 trillion different types of zygotes that are possible.
That's a LOT of variability, just from one pair of reproducing humans. Now, the differences in these 8 million gametes are slight. A couple of alleles, here and there. And, we are still talking about a human genome, so the entire set of genes still codes for a human; a gene for every trait. So, we are all fundamentally very similar to one another, sharing 99.9% of our genes. The ways we differ are actually pretty subtle... the color of hair, the slope of the nose, the bend of a finger. That's probably why other species seem so homogeneous to us until we really sit down and try to recognize these little personal differences.
That's the solution to Darwin's Dilemma about the source of variation.... hereditary particles don't blend together; they remain distinct and form new combinations from generation to generation, LARGELY AS A CONSEQUENCE OF INDEPENDENT ASSORTMENT, BUT ALSO AS A CONSEQUENCE OF CROSSING OVER, WHICH WE WILL DESCRIBE LATER. REALIZE THAT THE ARGUMENTS ABOVE ARE ONLY LOOKING AT THE EFFECTS OF INDEPENDENT ASSORTMENT. This huge amount of variation is not caused by the production of a new gene by mutation (although this does occur); this remarkable amount of variation is produced by recombination: the formation of new combinations of genes in gametes and offspring during sexual reproduction.
3. Model of Evolution circa 1905 (Rediscovery of Mendel's Laws)
Source of variation: Independent assortment of particulate hereditary particles during gamete formation.
Agents of change: NS.
Study Questions:
1. Diagram a cell, 2n=4, as it goes through Meiosis.
2. Explain how recombination creates an extraordinary amount of variation.
3. Explain how the movement of chromosomes in Meiosis I provides a mechanistic description of how Mendel's two principles work.
4. What are the costs and benfits of sexual and asexual reproduction?
5. How can male and female sexes be allocated in plants?
6. Distinguish between zygotic, gametic, and sporic meiosis.