 I.
A Brief History of LifeI.
A Brief History of Life
I.
A Brief History of LifeI.
A Brief History of Life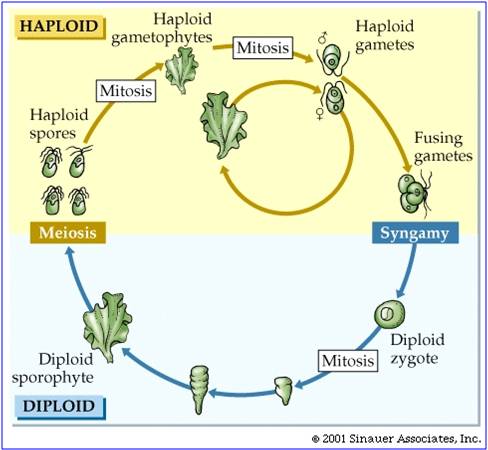 1.
Green Algal Ancestors
1.
Green Algal Ancestors
There is considerable evidence that plants evolved from green algal ancestors. They share the same type of chloroplasts (with chlorophyll a and b), and an intersting life history called "alternation of generation". In this life cycle, a haploid organism (the gametophyte) produces gametes by mitosis. These gametes fuse in a fertilization event, creating a diploid zygote. This diploid zygote divides (typically) mitotically to produce a multicellular diploid offspring. This organism produces haploid spores by meiosis. These spores develop, dividing mitotically to produce the next multicellular haploid generation. So, the ploidy level "alternates" across the generations. In green algae, the sporophyte and gametophyte stages may be "isomorphic" (look the same), like in sea lettuce (Ulva) show at right. In addition, green algae exhibit a range of gamete types, from isogametes that are double flagellated and the same size (as in Ulva) to oogamous species with double-flagellated sperm and immobile large eggs. All plants are oogamous, with large eggs and small sperm. In some gymnosperms and the angiosperms, the sperm have no flagella; in all other plants the sperm have flagella.
2. Colonization of Land: Environmental Differences
 The evolution of plants occurred as photosynthetic organisms acquired traits
that allowed for greater reproductive success on land - out of the ancestral
aquatic environment of algae. So, to understand how plants differ from algae,
and how and why plants evolved various characteristics, you must understand
how aquatic and terrestrial habitats differ. Algae and plants require several
essential nutrients: sunlight, water, and mineral nutrients (like nitrogen,
potassium, magnesium, and phosphorus, to name a few). They also require CO2
and oxygen, as all green algae and plants are photosynthetic aerobes, having
both chloroplasts (for photosynthesis that needs CO2) and mitochondria
(for aerobic respiration that needs O2). Aquatic habitats have ample
water (obviously). Carbon dioxide dissolves in water, so the concentration in
air and water will equilibrate over time. However, CO2 does not diffuse
quickly into or through water, so it may be less abundant locally. Likewise,
oxygen may be in short supply in aqueous habitats, largely because it does not
dissolve as readily. Although oxygen has a concentration of 21% in the atmosphere,
it is often about 7% or less in aquatic habitats, with more near the surface
and less at increasing water depths. In addition, water absorbs light energy
quickly. About 90% of the light energy is absorbed in the first 10m of water;
99% is absorbed by 50-100m. So, photosynthetic organisms must stay near the
top of a water column to get light and oxygen. Unfortunately, the process of
decomposition that recycles mineral nutrients tends to occur at the bottom of
a water column. As such, growth and photosynthesis of aquatic algae is usually
strongly limited by mineral nutrients - either phosphorus, nitrogen, or iron.
(This is why fertilizers running of into water can create huge algal blooms
- the factor previously limiting their growth has been provided, so they grow
like crazy). That is why the open ocean has little photosynthetic activity per
unit area; especially compared to shallower regions like continental margins
and shallow estuaries where upwelling and wave action bring nutrients to the
surface and feed local productivity. So, although aquatic habitats have lots
of water, terrestrial environments provide more sunlight, more oxygen, the same
amount of CO2 (but more equitably distributed and available), and
more mineral nutrients (which are eroding from the rocks or are being recycled
by decomposers and are present in the soil water). And in shallow-water environments
with a supply of nutrients, competition with other algae would become intense,
too.
The evolution of plants occurred as photosynthetic organisms acquired traits
that allowed for greater reproductive success on land - out of the ancestral
aquatic environment of algae. So, to understand how plants differ from algae,
and how and why plants evolved various characteristics, you must understand
how aquatic and terrestrial habitats differ. Algae and plants require several
essential nutrients: sunlight, water, and mineral nutrients (like nitrogen,
potassium, magnesium, and phosphorus, to name a few). They also require CO2
and oxygen, as all green algae and plants are photosynthetic aerobes, having
both chloroplasts (for photosynthesis that needs CO2) and mitochondria
(for aerobic respiration that needs O2). Aquatic habitats have ample
water (obviously). Carbon dioxide dissolves in water, so the concentration in
air and water will equilibrate over time. However, CO2 does not diffuse
quickly into or through water, so it may be less abundant locally. Likewise,
oxygen may be in short supply in aqueous habitats, largely because it does not
dissolve as readily. Although oxygen has a concentration of 21% in the atmosphere,
it is often about 7% or less in aquatic habitats, with more near the surface
and less at increasing water depths. In addition, water absorbs light energy
quickly. About 90% of the light energy is absorbed in the first 10m of water;
99% is absorbed by 50-100m. So, photosynthetic organisms must stay near the
top of a water column to get light and oxygen. Unfortunately, the process of
decomposition that recycles mineral nutrients tends to occur at the bottom of
a water column. As such, growth and photosynthesis of aquatic algae is usually
strongly limited by mineral nutrients - either phosphorus, nitrogen, or iron.
(This is why fertilizers running of into water can create huge algal blooms
- the factor previously limiting their growth has been provided, so they grow
like crazy). That is why the open ocean has little photosynthetic activity per
unit area; especially compared to shallower regions like continental margins
and shallow estuaries where upwelling and wave action bring nutrients to the
surface and feed local productivity. So, although aquatic habitats have lots
of water, terrestrial environments provide more sunlight, more oxygen, the same
amount of CO2 (but more equitably distributed and available), and
more mineral nutrients (which are eroding from the rocks or are being recycled
by decomposers and are present in the soil water). And in shallow-water environments
with a supply of nutrients, competition with other algae would become intense,
too.
In this context, land looks like a much better habitat for photosynthetic organisms than aquatic habitats. We can appreciate why selection might favor organisms that could colonize this environment. However, there is one significant hurdle - terrestrial environments are very very dry. Living tissue is mostly water; in order to survive in a terrestrial environment, living systems had to evolve mechanisms for reducing water loss. There are other problems that a deficit of water creates, too, like for reproduction. Algae and animals have swimming gametes; how can they reproduce on land? As we consider different groups of plants, we will focus on how plants evolved characteristics that solved these problems.
 3.
Plant Evolution: Adapting to Land
3.
Plant Evolution: Adapting to Land
The dominant plants on the planet today are the flowering plants (Angiosperms) and the conifers (a group of Gymnosperms). Both groups are fully terrestrial (except those that have recolonized water!), meaning that no stage of their life cycle requires an aquatic habitat to survive. They have a waxy cuticle on their leaves that reduces water loss, and they also have pores ("stomates") that close during dry conditions to reduce water loss still further. They have vascular tissue for the efficient transport of water from the roots to leaves ("xylem" tissue) and to transport sugars from leaves to the rest of the plant ("phloem" tissue). The xylem has cellulose and lignin in the cell walls, creating rigid tissue that can support the weight of the plant. In addition, these plants have non-motile sperm; the sperm is carried to the egg by the male gametophyte organism, the pollen grain. All of these characteristics are adaptations to life on land: reducing water loss, increasing the efficiency of water transport, and reducing dependancy on water as a necessary medium for gamete transport or bodily support. These adaptations did not evolve at once. Rather, through the evolution of different new plant groups, plants evolved and accumulated these adaptations. Today, although angiosperms dominate the plant kingdom, there are still representatives of more ancient groups. As we move through these groups, we will emphasize the major evolutionary innovations of each group.
4. Primitive Plants - the "non-tracheophytes"
 a.
Groups
a.
Groups
These groups are not a 'monophyletic' clade, but we will consider them together. They are the "liverworts", the "hornworts", and the "mosses". You are certainly aware of mosses, but liverworts are quite common, too, living on streambanks or on rocks in streams. Hornworts are a bit less common, but are also found in moist areas, including the bark of some trees.
b. Characteristics
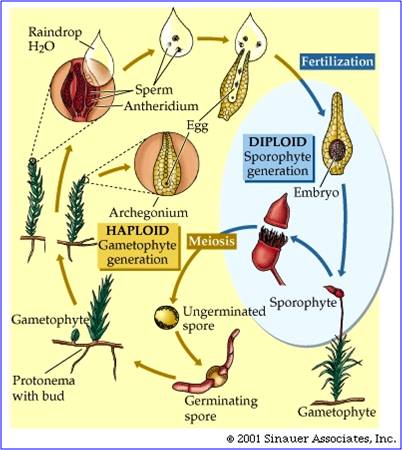
These groups are all non-vascular, meaning that they do not have xylem or phloem. As a consequence, water and sugar transport in the plants is from cell to cell and it is not very efficient. So, the plants must remain small and low to the ground - the source of their water. They also produce swimming sperm, and so must be covered by a thin film of water in order to reproduce. They do not have true leaves or roots (defined as having veins), and so water absorption is inefficient, too. In short, these plants are limited to living in very moist areas, and they must be covered by at least a thin film of water at some time during the year. These plants do have some adaptations for life on land that distinguish them from their algal ancestors, however. They secrete a thin oily cuticle that protects them a bit from water loss. In addition, hornworts and moss sporophytes have stomates - pores in their 'leaflets' to regulate gas exchange and water loss. Their life cycle is dominated by the haploid gametophyte generation; that means that when you are looking at a green moss plant or a liverwort, you are looking at a haploid gametophyte organism. The sporophyte organism is smaller, and is physiologically dependent on the haploid gametophyte parent. In moss, the sporophyte grows out of the archogonium (egg-bearing structure) on the top of the female moss gametophyte. It is brown, and receives the energy it needs to grow from the parent plant. The capsule that it produces is where meiosis occurs, reducing ploidy by 1/2 and producing the haploid spores that will each develop into new haploid gametophyte plants.
c. History
The evolutionary history of these plants is scant, because they do not produce thick, lignified cell walls that fossilize. However, scientists working in Argentina unearthed what they believe to be Liverwort spores that date to approximately 470 mya. This is the earliest fossil record of a terrestrial plant group.
5. Tracheophytes
 a.
Characteristics
a.
Characteristics
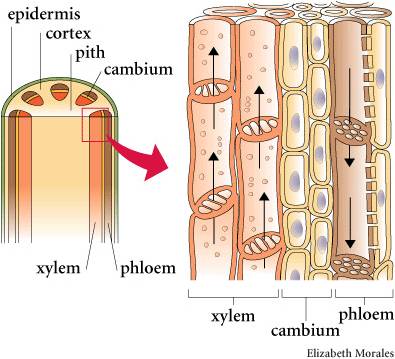
As the name denotes, these are the vascular plants that have true xylem and phloem tissue. Xylem tissue consists of cells that have died, leaving behind their highly lignified cell walls. The ends of these vessel elements have holes, allowing water to pass through like a straw. The evolution of xylem allows for efficient transport of water up a plant. This occurs through root pressure and evapotranspiration from leaves. By actively transporting salts into the xylem cavity, water follows by osmosis from living root tissue and water pressure 'pumps' water up the stem. At the other end of the xylem vessels, water is evaporating off leaves to the environment, creating a negative pressure that 'sucks' water up the stem. Because of the narrowness of the vessel elements, water also moves up by capillary action. All this means that plants could grow taller. This is advantageous for gathering light (the plant is not subject to shading by neighbors) and dispersing spores, but not too great for dispersing swimming sperm.
As such, it is not surprizing that the sporophyte generations becomes the dominant life history stage in tracheophytes. The gametophyte can't take advantage of larger size, because the plant needs to be covered by water for the sperm to swim. The sporophyte, however, benefits from being tall because the increased wind disperses spores farther.
b. Groups:
There are two major groups of tracheophytes, the Lycopodiophytes and the Euphyllophytes. These groups represent true evolutionary clades. They are distinguished by the types of leaves they have. The more primitive group, the Lycopodiophytes, have leaves with one unbranched vein. The Euphyllophytes (which means plants with true leaves), have leaves with many, often branching veins. The Lycopodiophyes include the clubmosses, spikemosses, and quillworts. The Euphyllophytes include the Monilophytes (ferns and kin), and the seed-bearing plants (Gymnosperms and angiosperms).
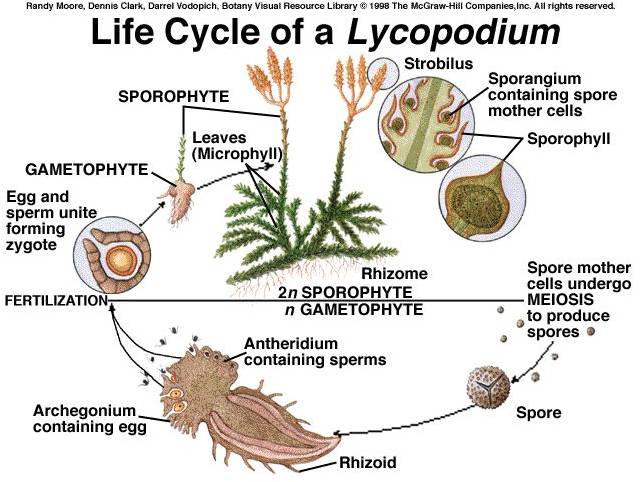 i.
Lycopodiophyta: These are the most primitive living vascular
plants, descended from a very diverse clade that dominated through the late
Silurian, Devonian, and Carboniferous Periods (400-300 mya) of the Paleozoic
Era. The presence of vascular tissue allowed their sporophytes to grow tall;
indeed, the 'arborescent' (tree-like) canopy of Devonian and Carboniferous forests
were clubmosses nearly 100 ft tall. Today, clubmosses are typically under a
foot in height, forming a lush carpet in the herb layer of moist forests. The
life cycle shows this inversion and dominance of the sporophyte. When you are
looking at a green clubmoss plant, you are seeing the diploid sporophyte. It
bears cone-like structures called strobili, and in those structures the sporangia
produce haploid spores by meiosis. The spores germinate, producing a free-living
gametophyte that lives in the soil. Archegonia and antheridia produce eggs and
sperm, respectively, by mitosis. The swimming sperm fertilize the eggs in the
archegonia, and the developing sporophyte grows out of its gametophyte parent,
as in the more primitive true mosses. However, this sporophyte is photosynthetically
active and grows to be a large, physiologically independent organism - it is
not dependent on its gametophyte parent as in true mosses.
i.
Lycopodiophyta: These are the most primitive living vascular
plants, descended from a very diverse clade that dominated through the late
Silurian, Devonian, and Carboniferous Periods (400-300 mya) of the Paleozoic
Era. The presence of vascular tissue allowed their sporophytes to grow tall;
indeed, the 'arborescent' (tree-like) canopy of Devonian and Carboniferous forests
were clubmosses nearly 100 ft tall. Today, clubmosses are typically under a
foot in height, forming a lush carpet in the herb layer of moist forests. The
life cycle shows this inversion and dominance of the sporophyte. When you are
looking at a green clubmoss plant, you are seeing the diploid sporophyte. It
bears cone-like structures called strobili, and in those structures the sporangia
produce haploid spores by meiosis. The spores germinate, producing a free-living
gametophyte that lives in the soil. Archegonia and antheridia produce eggs and
sperm, respectively, by mitosis. The swimming sperm fertilize the eggs in the
archegonia, and the developing sporophyte grows out of its gametophyte parent,
as in the more primitive true mosses. However, this sporophyte is photosynthetically
active and grows to be a large, physiologically independent organism - it is
not dependent on its gametophyte parent as in true mosses.
Study Questions:
1. How do liverworts differ from multicellular algae?
2. Draw the life cycle of a moss plant, showing ploidy levels and where the processes of mitosis and meiosis occur. Label gametes and spores.
3. What challenges face an aquatic algae as it tries to acquire light, gases, and nutrients?
4. To a photosynthetic organism, what are the benefits of colonizing land? What is the major "down side"?
5. Name two types of plant vascular tissue, describe what they do, and describe why vascular plants can be tall while non-vascular plants are short.
6. In clubmosses, the sporophyte is tall but the gametophyte is small. Explain why in terms of the reproductive cells they produce.
7. How do the Lycopodiophytes and Euphyllophytes differ?
8. Draw the life cycle of a club moss.