 I.
A Brief History of LifeI.
A Brief History of Life
I.
A Brief History of LifeI.
A Brief History of Life1. Green Algal Ancestors
2. Colonization of Land: Environmental Differences
3. Plant Evolution: Adapting to Land
4. Primitive Plants - the "non-tracheophytes"
5. Tracheophytes
a. Characteristics
b. Groups:
i. Lycopodiophyta:
ii. Monilophytes:
iii. Gymnosperms:
iv. Angiosperms: The angiosperms are the flowering plants; they are the most diverse group of plants on the planet today, represented by approximately 300,000 species and comprising 90% of all plant species. Angiosperms evolved early in the mesozoic, but radiated in the early Cretaceous Period - the last period of the Mesozoic Era. Although their diversity declined as a consequence of the meteor impact 65 mya (known as the K-T boundary because it is used to separate the Cretaceous ("K" - yeah, I know... but Cambrian got "C") of the Mesozoic from the Tertiary ("T") of the Cenozoic Era), they rebounded and dominated most terrestrial habitats. Like their sister group the gymnosperms, they have seeds and pollen. However, they also have two additional distinguishing derived characteristics - flowers and fruits.
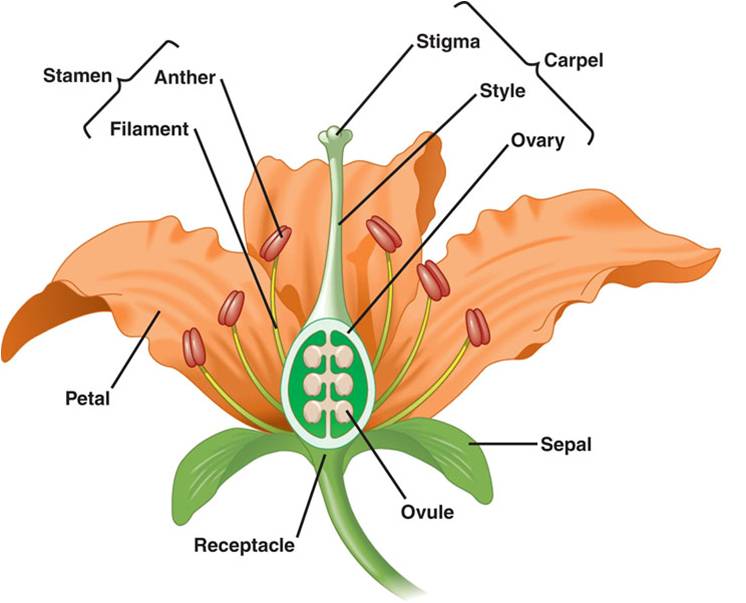 Flowers:
A flower is a stem-like structure that bears whorls of leaf-like appendages.
The lowest whorl on this reproductive "twig" is the calyx, composed
of leaf-like sepals that form a protective shealth surrounding the flower bud.
The second whorl of leaf-like appendages is the corolla, containing the leaf-like
petals. The third whorl contains the stamens. Each stamen has a filament that
bears an anther at the end. The anther is where meiosis occurs and haploid microspores
develop into pollen grains. At the center of the flower is the pistil, or carpel.
This is the female portion of the flower, consisting of the stigma (a sticky
surface that receives pollen), the style (a stalk connecting the stigma to the
ovary) and the ovary. The ovary has compartments that contain the ovules. The
structure of flowers can vary tremendously, and may lack carpels, petals, pistils,
or stamens. The number of the parts may vary, also, and 'composite' flowers
may contain thousands of flowers growing together as one big inflorescence -
with some flowers only bearing petals, and other flowers only bearing stamens
or ovaries. A daisy is a good example of a composite - the petals are actually
the product of single 'ray' flowers that each ONLY bear one petal. The yellow
center of a daisy is composed of hundreds of individual 'disk' flowers that
don't have petals but do have stamens and pistils.
Flowers:
A flower is a stem-like structure that bears whorls of leaf-like appendages.
The lowest whorl on this reproductive "twig" is the calyx, composed
of leaf-like sepals that form a protective shealth surrounding the flower bud.
The second whorl of leaf-like appendages is the corolla, containing the leaf-like
petals. The third whorl contains the stamens. Each stamen has a filament that
bears an anther at the end. The anther is where meiosis occurs and haploid microspores
develop into pollen grains. At the center of the flower is the pistil, or carpel.
This is the female portion of the flower, consisting of the stigma (a sticky
surface that receives pollen), the style (a stalk connecting the stigma to the
ovary) and the ovary. The ovary has compartments that contain the ovules. The
structure of flowers can vary tremendously, and may lack carpels, petals, pistils,
or stamens. The number of the parts may vary, also, and 'composite' flowers
may contain thousands of flowers growing together as one big inflorescence -
with some flowers only bearing petals, and other flowers only bearing stamens
or ovaries. A daisy is a good example of a composite - the petals are actually
the product of single 'ray' flowers that each ONLY bear one petal. The yellow
center of a daisy is composed of hundreds of individual 'disk' flowers that
don't have petals but do have stamens and pistils.
Flowers evolved to exploit animals for pollen transfer. Wind is capricious - the direction changes, the intensity changes, and even on a windy day only a small fraction of the pollen produced in a male cone will be lucky enough, just by chance, to travel to a female cone. For this reason, gymnosperms typically produce alot of pollen. By contrast, animals move through the environment non-randomly in search of food, shelter, and mates; and all animals can learn. So, by associating pollen with attractants (nectar, fragrance, and colorful floral displays) plants teach animals to visit flowers and increase the frequency of effective pollen transfer. Animals visiting an attractive flower are rewarded with food and quickly learn the association; they then travel in "bee-lines" from one flower to another seeking food, transfering pollen non-randomly and efficiently. This would be particularly useful for plants in the understory of a forest, where wind is minimal. The first angiosperms may have been shrubby, understory species. Animal pollination would also be very adaptive in tropical rainforests or deserts, where members of a single plant species may be very far away (decreasing the chances that a random procees like wind pollination would effectively find a 'mate').
As some angiosperms evolved into taller forms where wind could be used effectively, some were selected to use wind as a pollen-dispersal mechanism - with correlated reductions in flower size. So, wind-pollinated flowering trees (oaks, maples, etc.) and wind-pollinated field plants (open fields are windy environments, too) like grasses often have very small flowers with no colorful petals or sepals. Because they are not attracting animals for pollen dispersal, selection favors organisms that do not invest alot of energy in these structures.
 Plants
can increase the efficiency of pollen transfer even further by limiting flower
visitation to one type of pollinator. A specialized pollinator that visits just
one type of flower is more likely to transfer pollen between plants of the same
species. Likewise, flowers visited by just one type of pollinator are likely
to have nectar present when that pollintor visits. So, "pollination syndromes"
have evolved, representing flower traits that are associated with a particular
type of pollinator. For example, many flowers are red and tubelike. These flowers
are often pollinated by hummingbirds, which see into the red end of the spectrum
better than most insects and have a long beak that can access the nectar at
the base of the flower. Butterfly flowers are often composites (like daisies,
and zinnias), or are clustered in groups (butterfly bush). This allows the butterfly
to land on a cluster and probe many individual flowers at a time with their
long proboscis. Moth and bat pollinated flowers open at night. They are usually
white and very fragrant, attracting their pollinators with scent. Some plants
like skunk cabbage produce flowers that smell like rotting flesh. This attracts
flies that act as pollinators. Some of the most elaborate contrivances by which
plants limit pollination to a single species of pollinator are seen in the orchid
family. Some orchids produce a scent that mimics the sexual pheromones of females
of specific species of solitary wasps. When males perceive the scent, they fly
up the concentration gradient to the flower. In some species, the flower also
mimics the shape and coloration of the female wasp! The deception is so good
that the male attempts to mate with the flower. It gets dusted with pollen,
and flies off. While this may be frustrating for the wasp (who knows), it can't
NOT fly down the next pheromone trail it encounters or it won't mate. So, if
it flies down another trail and is tricked by another plant, pollen is transferred.
The bucket orchid is another strange plant. Two petals have fused into a bucket
that collect water. Pollinators fall in the bucket, and the only exit is through
a tight squeezeway where they are dusted with pollen.
Plants
can increase the efficiency of pollen transfer even further by limiting flower
visitation to one type of pollinator. A specialized pollinator that visits just
one type of flower is more likely to transfer pollen between plants of the same
species. Likewise, flowers visited by just one type of pollinator are likely
to have nectar present when that pollintor visits. So, "pollination syndromes"
have evolved, representing flower traits that are associated with a particular
type of pollinator. For example, many flowers are red and tubelike. These flowers
are often pollinated by hummingbirds, which see into the red end of the spectrum
better than most insects and have a long beak that can access the nectar at
the base of the flower. Butterfly flowers are often composites (like daisies,
and zinnias), or are clustered in groups (butterfly bush). This allows the butterfly
to land on a cluster and probe many individual flowers at a time with their
long proboscis. Moth and bat pollinated flowers open at night. They are usually
white and very fragrant, attracting their pollinators with scent. Some plants
like skunk cabbage produce flowers that smell like rotting flesh. This attracts
flies that act as pollinators. Some of the most elaborate contrivances by which
plants limit pollination to a single species of pollinator are seen in the orchid
family. Some orchids produce a scent that mimics the sexual pheromones of females
of specific species of solitary wasps. When males perceive the scent, they fly
up the concentration gradient to the flower. In some species, the flower also
mimics the shape and coloration of the female wasp! The deception is so good
that the male attempts to mate with the flower. It gets dusted with pollen,
and flies off. While this may be frustrating for the wasp (who knows), it can't
NOT fly down the next pheromone trail it encounters or it won't mate. So, if
it flies down another trail and is tricked by another plant, pollen is transferred.
The bucket orchid is another strange plant. Two petals have fused into a bucket
that collect water. Pollinators fall in the bucket, and the only exit is through
a tight squeezeway where they are dusted with pollen.
Fruits:Many angiosperms also evolved mechanisms to use animals to disperse their seeds. The ancestral condition within the group is probably the production of fleshy, ovary tissue that surrounds the carpels. Like nectar, this fleshy fruit tissue serves as a bribe, encouraging animals to eat the fruits and thereby disperse the seeds. And since animals tend to show a preference for a given habitat type, the seeds get transferred to another place with similar environmental conditions (rather than being strewn haphazardly across the environment by wind). Other fruits are dry - like acorns, grains, burrs, and some beans. If these are small, like in grasses, or have wings like maple samaras, or have feathery structures like milkweed, then the fruit can be dispersed by the wind. Some, like cocklebur, burdock, and goatshead, evolved hooks on their seed pods that could tangle in mammal fur. Some fruits are pea-pod like, and when the pod dries, the seam bursts open and the seeds are "shot out". This is called 'ballistic' dispersal. So, regardless of whether the fruit is animal or wind dispersed, plants have evolve many different ways to get their seeds dispersed and away from the competitive environment beneath the parent plant.
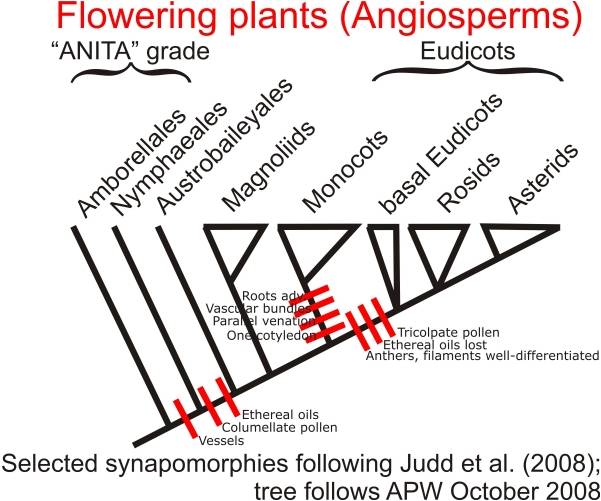 Diversity:
The 300,000 angiosperm species have radiated into such a wide variety of shapes
and behaviors that the evolutionary relationships among major groups (which
each contain lots of diversity) is often difficult. However, several groups
do fall out, especially with molecular analyses. Two ancient, basal groups are
the Amborellales ( a small group of a few very primitive species) and the Nymphaeales
(water lilies and their kin). Another ancient group is the Magnoliales, including
the Magnolias and the laurels. Two other large and diverse groups are the monocots
(including grasses, palms, lilies, and orchids), and the dicots (including 70%
of all plants, from roses to oaks, from buttercups to maples). Monocots have
one embryonic leaf (cotyledon), while dicots have two. Also, monocots have leaves
with parallel veins, while dicots usually have leaves with a network of radiating
veins. Monocots typically have flower parts in multiples of 3's, while dicots
have flower parts in multiples of 4's or 5's.
Diversity:
The 300,000 angiosperm species have radiated into such a wide variety of shapes
and behaviors that the evolutionary relationships among major groups (which
each contain lots of diversity) is often difficult. However, several groups
do fall out, especially with molecular analyses. Two ancient, basal groups are
the Amborellales ( a small group of a few very primitive species) and the Nymphaeales
(water lilies and their kin). Another ancient group is the Magnoliales, including
the Magnolias and the laurels. Two other large and diverse groups are the monocots
(including grasses, palms, lilies, and orchids), and the dicots (including 70%
of all plants, from roses to oaks, from buttercups to maples). Monocots have
one embryonic leaf (cotyledon), while dicots have two. Also, monocots have leaves
with parallel veins, while dicots usually have leaves with a network of radiating
veins. Monocots typically have flower parts in multiples of 3's, while dicots
have flower parts in multiples of 4's or 5's.
Life Cycle: The generalized life cycle of angiosperms is similar to gymnosperms. The diploid sporophyte produces two types of spores. In angiosperms, however, these can be born by the same structure (perfect hermaphroditic flowers), or different flowers on the same plant (monoecious) or on different plants (dioecious).
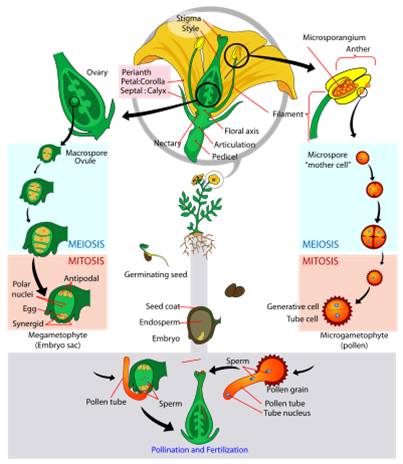 Pollen
lands on the stigma, and one tube cell secretes enzymes that dig a "pollen
tube" through the style to the ovule. It is followed down the tube by two
sperm cells. When they reach the opening of the ovule, one sperm cell fertilizes
the egg and forms the diploid zygote. The other sperm cell fuses with the TWO
polar nuclei in the middle of the ovule, forming a triploid nucleus. This "double
fertilization" is unique to the angiosperms. The triploid nucleus divides,
forming triploid 'endosperm' tissue within the ovule that surrounds the developing
diploid enbryo and provides nutrition. So again, the seed contains three layers
- the ovule of the original diploid sporophyte, the triploid endosperm, and
the diploid embryo.
Pollen
lands on the stigma, and one tube cell secretes enzymes that dig a "pollen
tube" through the style to the ovule. It is followed down the tube by two
sperm cells. When they reach the opening of the ovule, one sperm cell fertilizes
the egg and forms the diploid zygote. The other sperm cell fuses with the TWO
polar nuclei in the middle of the ovule, forming a triploid nucleus. This "double
fertilization" is unique to the angiosperms. The triploid nucleus divides,
forming triploid 'endosperm' tissue within the ovule that surrounds the developing
diploid enbryo and provides nutrition. So again, the seed contains three layers
- the ovule of the original diploid sporophyte, the triploid endosperm, and
the diploid embryo.
6. Summary:
Over the course of plant evolution, organisms evolve characteristics that provided a selective advantage in progessively more desiccating environments, allowing plants to colonize new habitats unoccupied by other plants. The non-tracehophytes evolved a cuticle, and could colonize land and harvest more sunlight than their algal ancestors. Club mosses evolved vascular tissue and a dominant sporophyte, allowing them to grow tall and gain a competitive advantage over smaller non-tracheophytes. Ferns evolved more complex leaves, improving the transport of water and sugars into and out of leaves. The evolution of seeds, the reduction of the the gametophyte, and the elimination of swimming sperm allowed seed ferns and their descendants - the gymnosperms and the angiosperms - to colonize upland areas away from water. Conifers still dominate in the boreal forests of high latitudes; however, the angiosperms dominate in almost all other terrestrial habitats. Their innovations of flowers and fruits exploit animals in the environment, and improve the efficiency of pollen transport and seed dispersal. The evolution of plants is an interesting story of adapting to life on land, to take advantage of the abundant sunlight, oxygen, and mineral nutrients found in this habitat. The are an extraordinarily important, interesting, diverse, and beautiful group of living organisms.
Study Questions:
1. Describe the parts of a flower.
2. How do showy flowers increase the efficiency of pollen transfer?
3. Describe three pollination syndromes.
4. Where doe the angiosperms that use wind dispersal live? Why have they evolved small flowers?
5. What is the adaptive advantage of fleshy fruit production?