 III.
The Prokaryote Domains: Bacteria and ArchaeaIII.
The Prokaryote Domains: Bacteria and Archaea
III.
The Prokaryote Domains: Bacteria and ArchaeaIII.
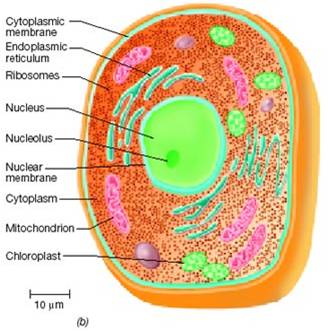
The Prokaryote Domains: Bacteria and ArchaeaThe eukaryotes have been evolving for at least 2.0 billion years. As you might expect (as was argued for the prokaryotes groups), there has been considerable divergence within this group, producing a dramatic range of genetic and metabolic variation. And of course, the multicellular eukaryotes evolved from protistan ancestors - so multicellular eukaryotes (plants, fungi, and animals) are included within this clade. So, the focus will be on the origin of the clade (Eukarya) and the diversity within this clade. We will pay particular attention to the sister groups of the multicellular "kingdoms" that we will consider in greater detail in the remaining lectures. Obviously, the features that distinguish eukaryotes from archaean ancestors are the presence of membrane-bound organelles (mitochondria and chloroplasts) and a nucleus. So, any hypothesis of origin must explain how these defining structures came into existence.
1. Origin of Organelles
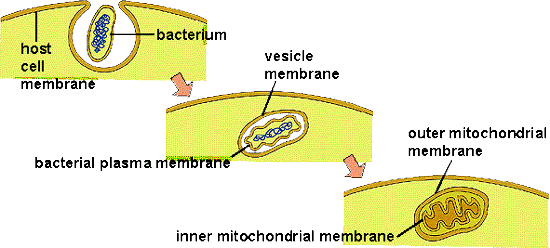
The origin of eukaryotic cells is described by the Endosymbiotic Theory, popularized by Lynn Margulis in the 1960's. It states that organelles are descended from free-living bacterial cells that were absorbed by - or parasitized - archaeans. The evidence for this idea is very strong. Both mitochondria and chloroplasts have double membranes. The outer membranes are similar to the plasma membrane of eukaryotes and archaeans. The inner membrane has certain attributes of bacteria. If a bacterium were absorbed through the process of endocytosis, the bacterium would be surrounded by the membrane of the host's vacuole - creating a double membrane. In addition, both types of organelles have their own DNA, which is circular like bacterial DNA. Both mitochondria and chloroplasts divide by fission, independent of the mitotic division of the eukaryotic cell they inhabit. Over the course of evolutionary history, genes have been exchanged between the organelles and the nucleus, or have been eliminated in the organelle because they also exist in the nucleus. For these reasons, the organelles are functionally dependent on the eukaryotic cell and can no longer live independently for very long.
Additional similarities link these organelles to specific bacteria. Mitochondria not only have their own DNA, but they also produce their own proteins using these DNA recipes. (That should be no surprize!) Protein synthesis occurs on ribosomes within the mitochondria, and their ribosomes are more similar to bacterial ribosomes than to the ribosomes in the cytoplasm of their eukaryotic host cells (produced by the nuclear DNA). Likewise, the genetic code used by mitochondria has a few differences from the universal genetic code; differences also seen in the proteobacteria. Genetic analyses suggest that mitochondria are most closely related to Rickettsia proteobacteria. Rickettsia does not causes "rickets" (a vitamin-D deficiency); rather, Rickettsia bacteria cause a variety of human diseases like many forms of tick-borne fevers (Rocky Mountain spotted fever) and typhus. In addition to having a similar genome to mitochondria, these bacteria are obligate intracellular parasites - so they move into eukaryotic cells and become "organelle-like". In addition, Rickettsia produce ATP in the same way that mitochondria do. So, it is possible that archaeans absorbed bacteria and used them for ATP production, or that the bacteria infected the archaeans to consume their sugars, or some combination of these scenarios. In any case, the bacteria and archaean ultimately benefitted from the interaction, forming a mutualistic or "symbiotic" relationship.
 |
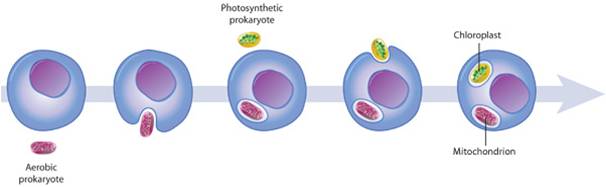 |
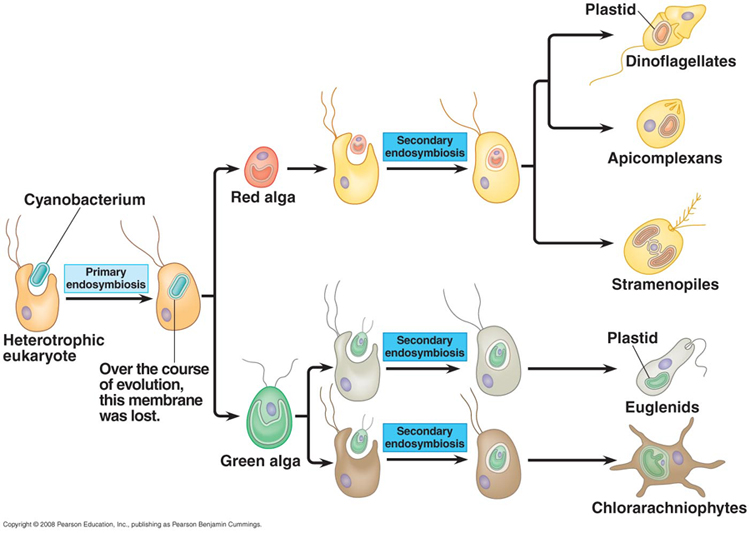
Chloroplasts have an independent, endosymbiotic origin; they are genetically similar to a different group of bacteria called cyanobacteria. Some have a double membrane like mitochondria. However, in some protistan groups, the chloroplasts have 3 or 4 membranes, suggesting multiple rounds of endosymbiosis. So, the current model of chloroplast ancestry suggests a single origin of photosynthetic red and green algae. Green algae were then absorbed (secondary endosymbiosis) by other unrelated protists, creating the euglenids and chlorarachniophytes. These organisms are not related directly through nuclear genetic analysis, but they have very similar, multi-membraned chloroplasts - suggesting independent endocytosis of green algae. A similar process may explain the chloroplast similarity in the stramenopiles, apicomplexans and dinoflagellates - independent endocytosis of a red algae could explain their dissimilar nuclear DNA and their similar, multi-membraned chloroplasts.
 2.
Origin of the Nuclear Membrane
2.
Origin of the Nuclear Membrane
There are several hypotheses regarding the evolution of the nuclear membrane. Like organelles, the nuclear membrane is a double membrane - suggesting that it may have evolved from endosymbiosis. Because of the greater genetic similarity between archaeans and eukaryotes, the hypothesis is that an archaean cell was engulfed by another cell (archaean or proteobacterian). Ultimately, perhaps through gene transfer or decay, the genes in the internalized cell came to dominate the metabolic activity of the cell. Another hypothesis is that the nuclear membrane formed by invaginations of the plasma membrane, encapsulating the chromosomes and isolating them from the more enzymatically active region of the cell (cytoplasm).
There are several groups of eukarya; most of them contain only single-celled representatives.
 1.
Diplomonada and Parabasalida
1.
Diplomonada and Parabasalida
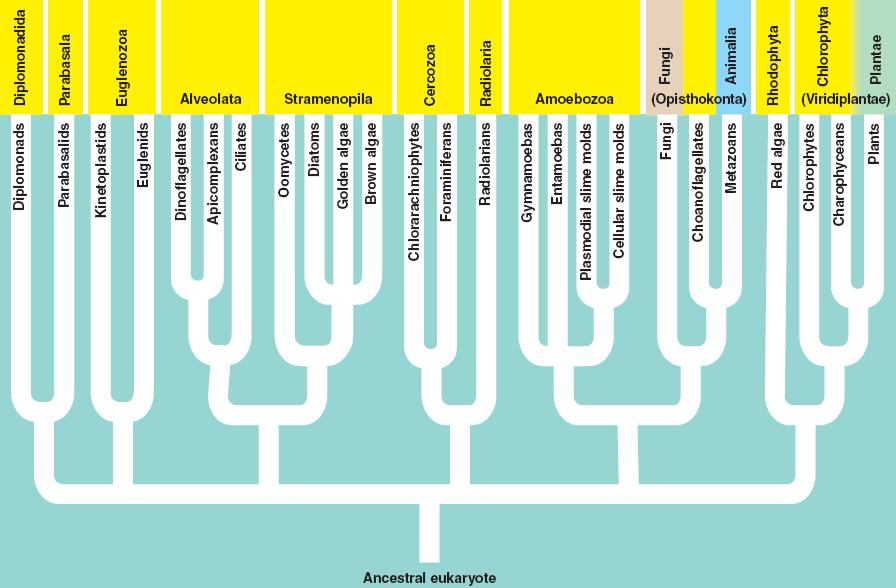
These two groups are unusual in having very reduced (even absent) mitochondria. 'Reduced' means that they do not perform all of the functions of mitochondria found in other eukaryotes. Although some scientisits initially thought that these might be transitional forms between archaeans and eukaryotes, the thought now is that these groups have lost mitochondrial function because they have adapted to anaerobic environments. Without oxygen present, the mitochondria becomes a non-functional, vestigial organelle. Giardia is a diplomonad (two equal nuclei) and Trichomonas is a Parabasalid. Giardia is an intestinal parasite of vertebrates, and can infect the human gut where it causes acute diarrhea, gas, bloating, and dyehydration and weight loss. Trichomonas vaginalis infects the vagina and urogenital tract; it is a common sexually transmitted disease that usually causes no symptoms in males but does cause itching, discomfort, and discharge in females. This infection is called trichomoniasis or 'trich'.
2. Euglenozoa
This group includes the euglenids and the kinetoplastids - two groups in which the cells have a supportive rod in the flagella. Sleeping sickness is caused by members of this group, the Trypanosomes. The euglenids are unusual in being heterophototrophs. They can synthesize their own sugars through photosynthesis, but they can also prey on small prey items. Their chloroplasts have three membranes and are believed to have resulted from secondary endosymbiosis of green algae.
 3.
Alveolata
3.
Alveolata
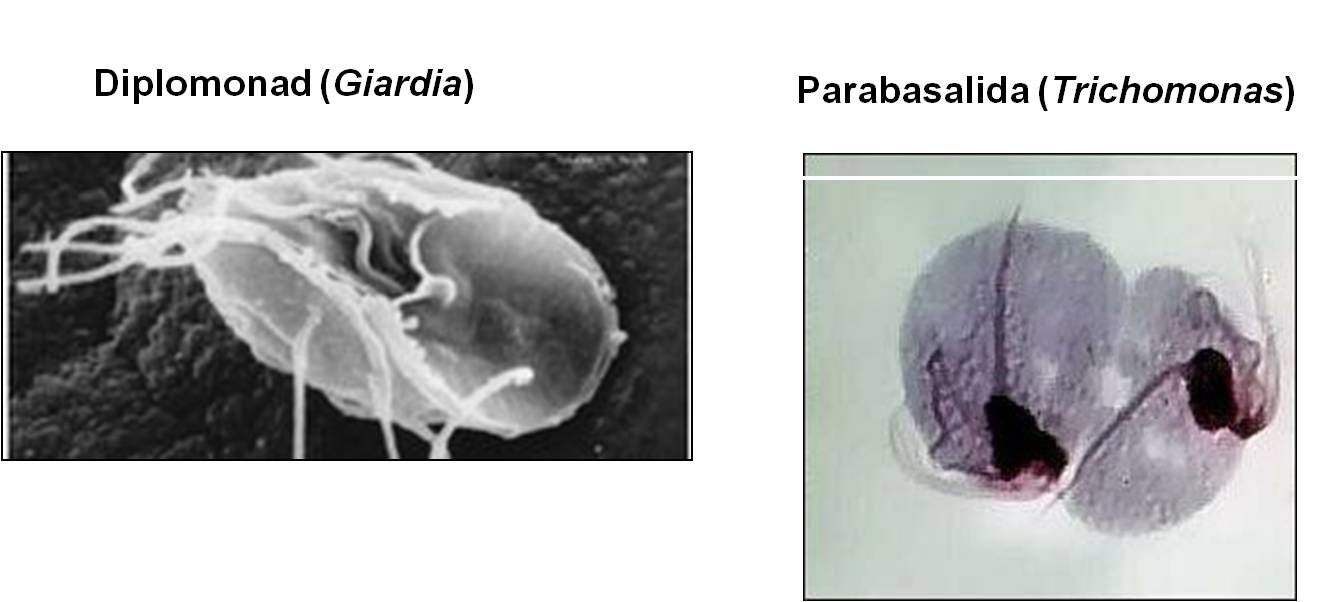
This group of diverse organisms have membrane-bound vesicles nested beneath the plasma membrane. It includes a wide variety of photosynthetic, parasitic, and predatory organisms, including ciliates like Paramecium, Dinoflagellates (cause of 'red tides'), and Apicomplexans like the malarial parasites in the genus Plasmodium.
4. Stramenopiles
This is another diverse group of photosynthetic and heterotrophic organisms, characterized by their two flagella being 'hairy'. Diatoms, golden algae, and brown algae are photosynthetic members of this group. Kelp is a huge multicellular brown algae. They have chloroplasts with four membranes that are thought to have arisen by secondary symbiosis of a red algae.
5. Rhizaria
These sister taxa have thin, threadlike pseudopods that they extend through a shell or 'test'. The Foraminiferans, belong to the Cercozoa group, often harbor symbiotic algae and feed off the sugars these algae produce. By contrast, most radiolarians are predatory, feeding on bacteria and protists that get 'stuck' to these sticky cellular extensions. Deep sedimentary deposits of fossil foraminiferan and radiolarian tests have been found, and are often used as bioindicators that specify strata of a particular age.
6. Amoebozoa
This clade includes organisms with broad pseudopods that are used for locomotion and prey capture. Most amoebozoans are free-living, living in moist soil or freshwater environments. However, the Entamoebas are parasitic, infecting the digestive tract (where they cause amoebic dysentery) or the brain (amoebic meningitis) - both of which can be fatal. Slime molds are also included in this clade. Although they were once thought to be fungal, they do have an amoeboid stage that confirms their genetic relationship to this group. Some slime molds have a strange life cycle in which free-living, independent amoebas aggregate and form a spore-producing structure. Some cells in the mass dry out and become a rigid stalk, while other cells migrate to the end of the stalk and develop into encapsulated spores. New amoebae 'hatch" from the dispersed spores when conditions are favorable. This is an interesting case where protist cells cooperate to form a multicelluar aggregate with the rudiments of cell specialization. However, this is not the group from which multicellular animals or fungi evolved.
 7.
Opisthokonta
7.
Opisthokonta
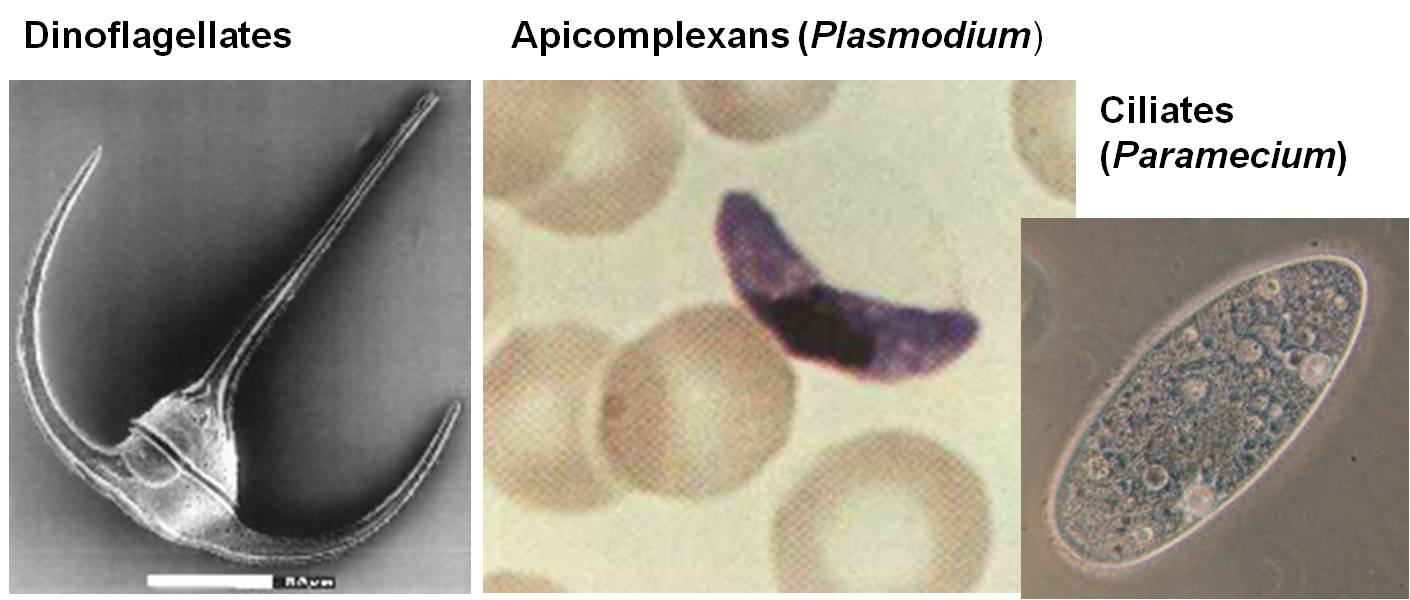
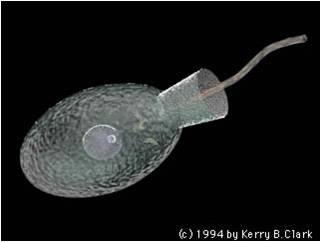
This clade includes the fungi and animals, and the flagellated sister taxa that bear the stamp of their close phylogenetic relatedness. Other groups have flagella, like the diplomonads and the euglenozoa - but those groups have two flagella and they are at the 'front' of the organism, pulling the organism along surfaces or through a liquid medium. The Opisthokonts, however, have a single, posterior flagellum at some point in the life cycle of the organism. In the animals, this is represented by sperm cells. most fungi do not have flagellated cells, except for the most promitive group, the chytrids. This morphological characteristic is the only morphological trait that links these groups, but their genetic similarity testifies to their common ancestry.
An important opisthokont group besides the fungi and animals are the Choanoflagellates. This group is considered to be the "sister group" to the animals, and may represent what the ancestral animals evolved from. The Choanoflagellates are distinguished by a ring of small projections that ring the flagellum like a collar. These microvilli act like a sieve. The motion of the flagellum wafts bacteria into the ring of microvilli, where bacteria are trapped and then absorbed by phagocytosis. There are over 100 species of choanoflagellates; some are solitary, but some form colonies. Their predation of bacteria provides a valuable link in marine food webs. As we will see when we get to animals, the most primitive animals - the sponges - have cells that are very similar to these choanoflagellates.
 8.
Archaeplastida
8.
Archaeplastida
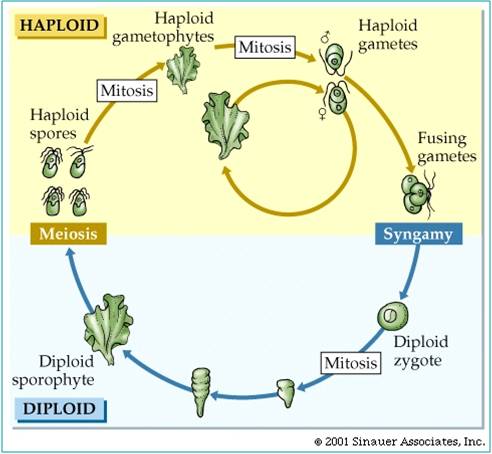
The last eukaryotes clade contains red algae, green algae, and land plants. Red algae have chloroplasts that contain chlorophyll a like cyanobacteria; green algae and plants contain chloroplasts with chlorophyll a and chlorophyll b. The red algae are largely multicellular, colonial organisms best represented by red seaweeds. Green algae are a diverse group, including unicellular, colonial, and multicellular species like Ulva and Enteromorpha (green seaweeds), Volvox, and Spirogyra. A key shared derived character that is seen in the Archaeplastida is "alternation of generation". Consider a haploid organism about to reproduce. This organism produces gametes (egg and/or sperm) by mitosis. These gametes fuse to form a diploid zygote, which developes into the diploid offspring. The diploid organism produces haploid spores by meiosis. These spores do not fertilize one another. Rather, they each divide by mitosis and develop into separate haploid organisms. So, the ploidy level alternates each generation. All plants share this life cycle, so we will see it often during the plant diversity lectures.
Study Questions:
1. What characteristics distinguish the Domain Eukarya?
2. Describe the endosymbiotic theory of organelle origin, and provide evidence for this theory.
3. Describe two hypotheses regarding the evolution of the nuclear membrane.
4. What characteristics do the Opisthokonts share, and what three groups are included in this clade?
5. What characteristics do the Archaeplastida share, and what three groups are included in this clade?
6. What is 'secondary endosymbiosis' and what groups nay have acquired their chloroplasts through this process?